АгроСборник.Ру
С.Ф. Коваль, В.П. Шаманин "Растение в опыте"
1.1. Выше адаптация рассматривалась в плане системной организации растения. Одновременно с этим, возможна классификация адаптивных реакций в соответствии с временной динамикой развития неблагоприятного воздействия. Возникновению неблагоприятного сезонного или погодного комплекса предшествуют с высокой достоверностью некоторые нейтральные события: зиме - сокращение длины дня, а лету - увеличение его продолжительности и т.д. Таким образом, сокращение длины дня является для растения сигналом приближающейся зимы. Озимые сорта при осеннем посеве генетически запрограммированы переходить к генеративному развитию только после получения определенного числа дней с отрицательной температурой. Повышение температуры почвы и воздуха после экспозиции холодом означает для озимого растения начало весны и является сигналом к возобновлению вегетации. Но на юге возможен временный возврат тепла зимой, при котором озимые растения возобновляют вегетацию и погибают при новом понижении температуры. Защитой от преждевременного возобновления вегетации в зимние оттепели служит реакция этих растений на короткий зимний день, который также является сигналом продолжающейся зимы. Осенью озимые растения реагируют на короткий день торможением роста, накапливают в тканях неизрасходованные метаболиты и благодаря этому закаляются в преддверии наступающей зимы. Показано [Коваль, Ахмедов, 1989], что рекордно высокая зимостойкость наблюдается только у тех сортов озимой пшеницы, которые наиболее чувствительны к длине дня. Таким образом, реакции стадийного развития (яровизация и фотопериодизм) имеют целью определение характера сезона и синхронизацию развития растения с ходом климатического годового цикла. К этому же кругу явлений следует отнести и глубокий зимний покой деревьев и кустарников, а в зоологии - диапаузу насекомых, зимнюю спячку некоторых млекопитающих и широко распространенную у животных синхронизацию периода гона с длиной дня. До начала почвенной засухи (главным фактором которой является повышенная дневная температура и круглосуточный водный дефицит) в течение ряда дней или даже недель погода характеризуется низкой влажностью воздуха, прозрачной атмосферой и большой интенсивностью солнечного света, обогащенного ультрафиолетом. Этот комплекс факторов сам по себе не вызывает нарушения физиологических процессов, однако приводит к торможению роста. А последнее, как отмечалось выше, является эффективным путем формирования организменной устойчивости. Поскольку в подобных случаях адаптация предваряет прямое повреждающее воздействие, мы выделяем эти реакции в группу заблаговременных.
Еще одни пример заблаговременных реакций представляют нам общеизвестные нормы реакции растений на микроусловия произрастания в пределах ареала вида. Распределение семян в большинстве случаев носит случайный характер и различные биотипы могут случайно распределяться по ландшафту, занимая экологические ниши на северных или южных склонах, в долине или на возвышенности. Сохранение вида в значительной степени определяется гетерогенностью популяции семян, выживанием в каждой нише местообитания наиболее адекватных генотипов, но важную роль играет и норма реакции генотипа. Так, на южных склонах повышенная дневная температура вместе с хорошей освещенностью имитируют природный комплекс условий, предшествующий засухе. Эти условия сдерживают ростовые процессы и обеспечивают формирование повышенной (по сравнению с растениями северных склонов) устойчивости еще до иссушения почвы. К группе заблаговременных реакций могут быть отнесены не только явления фото- и термопериодизма, но и процессы морфогенеза, изменяющие через регуляцию роста габитус растения и положение его органов относительно поверхности почвы. Рассмотрим последнюю на примере регуляции глубины залегания в почве узлов кущения злаков. Оптимальная глубина залегания в почве узлов кущения как у многолетних, гак и у однолетних злаков должна удовлетворять достаточной защите их от иссушения летом и от вымерзания зимой и, в то же время, не препятствовать выходу на поверхность молодых побегов, образующихся из почек узла. В реальной обстановке оптимальное положение узла кущения зависит от механического состава почвы, ее плотности, влажности, температуры освещенности и ряда других факторов. Главным фактором остановки роста эпикотиля является свет. Если семена высеять в стеклянные сосуды вплотную к его стенке, то проростки получают освещение под землей с первого дня прорастания. Эпикотиль при этом не развивается, и узел кущения формируется слитым со щитком, как это бывает при прорастании семян на поверхности почвы. В процессе формирования всходов из заглубленных в почву семян стеблевой узел первого листа продвигается внутри колеоптиля к поверхности почвы в результате роста первого междоузлия (эпикотиля). При достижении узлом оптимальной глубины рост эпикотиля прекращается. Междоузлия стебля вышележащих фитомеров не растут, и их стеблевые узлы со спящими почками сливаются с первым узлом, образуя будущий узел кущения (рис. 2.2). Таким образом, оптимальное положение в почве узла кущения зависит от своевременного прекращения роста эпикотиля, которое должно согласовываться с величиной многих параметров среды, имеющих различную физическую природу.
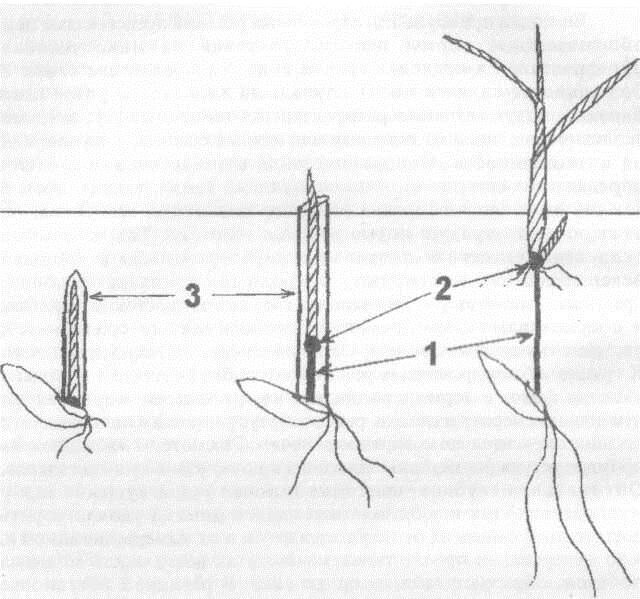
Рис. 2.2. Формирование узла кущения у злаков. 1 - эникотиль; 2 - узел кущения; 3 - колеоптиль
Наши исследования [Коваль, 1972; 1973а] показали, что сигналом к прекращению роста эпикотиля является повышение концентрации кислорода в тканях проростка. Кислород в тканях расходуется в процессе дыхания, а его поступление из воздуха уменьшено за счет сопротивления диффузии газов через почву и ткани колеоптиля. Чем толще покрывающий слой почвы и больше ее плотность и чем выше тургор тканей, тем больше сопротивление диффузии и тем ниже концентрация кислорода в тканях. С приближением листа к освещенной поверхности концентрация кислорода возрастает, сначала благодаря частичному подавлению дыхания светом, а затем - за счет фотосинтеза. Низкая концентрация кислорода в тканях проростков установлена нами прямым полярографическим определением кислорода [Коваль, 19736], а его роль в торможении роста - нарушением диффузионного сопротивления покровных тканей [Коваль, 1969].
В итоге, все внешние факторы, влияющие на положение узла кущения относительно поверхности почвы, действуют посредством усиления или ослабления аэробности внутренней газовой среды растения (воздуха межклетников и газа, растворенного во внутриклеточной воде). Этим достигается сопоставление и объединение информации (различной по физической природе носителей) в виде единого сигнала, по которому генетически запрограммированные системы определяют оптимальный момент прекращения роста эпикотиля. Интеграция в едином сигнале многих параметров среды уменьшает вероятность ошибки, которая могла бы иметь для растения роковые последствия. Как мы видим, физиологогенетическая программа заблаговременной реакции включается по внешнему сигналу, который сам по себе не является отрицательным экологическим фактором, но в течение многих поколений его возникновение коррелировало с последующим ухудшением экологической обстановки. В процессе эволюции происходил отбор генотипов, отвечающих на появление фактора-сигнала адаптивными реакциями, опережающими возникновение стрессовых ситуаций. предсгавленис об опережающем отражении событий в общем плане и в применении к животным и человеку разрабатывались в трудах П.К. Анохина 11978], но применительно к растению этот принцип ранее был изложен только в одном коротком сообщении [Коваль, 1974-а]. В рассмотренном нами примере регуляции положения узлов кущения мы имеем дело с крайне интересной проблемой распознавания образа биологическим объектом. В отношении растительного организма эта проблема, насколько нам известно, не исследована, но в биохимии и в молекулярной биологии хорошо известны случаи взаимного распознавания молекул во многих реакциях. Анализируя проблему распознавания образа в биологии, Кастлер [1968] отмечает, что во взаимодействии фермента с субстратом последний распознается не по строению всей молекулы, а только по некоторой характерной ее части, называемой сигнатурой. У различных молекул характерные части могут быть одинаковыми, и тогда возникают ошибки распознавания (по причине двусмысленности сигнатуры). В качестве примера можно привести реакцию присоединения к гемоглобину молекулы угарного газа вместо кислорода. Для более точного распознавания возможна идентификация субстрата не по одному, а по нескольким характерным участкам (сигнатурам). Однако Кастлер не касается вопроса об обобщении информации сигнатур, представленных количественными признаками, и о распознавании образа в случае, когда сигнатуры противоречат друг другу. Видимо, и в заблаговременных реакциях используется сигнатурный принцип распознавания экологической обстановки.
Каждый параметр среды, влияющий на положение узлов кущения в почве, может с некоторой вероятностью дать ошибочную информацию. Можно предположить, что во многих случаях интегральным сигналом, в котором объединяются различные сигнатуры, может быть концентрация гормонов и их ингибиторов в растении. Учет многих параметров различной физической природы снижает общую ошибку распознавания. Но для интеграции первичных сигналов необходимо сложение их, а значит - перевод с определенным для каждого из них коэффициентом в единый сигнал. При моделировании тех или иных условий в опыте следует учитывать возможность заблаговременных реакций растения. Исследователь может просто не обратить внимания на фактор, не имеющий прямого отношения к основным процессам жизнедеятельности, который, однако, является для растения сигналом появления определенной экологической ситуации. Возникшая в результате этого реакция растения кажется неожиданной и часто служит предметом длительных размышлений на тему «почему это случилось?»
При выращивании в регулируемых условиях среды отсутствие некоторых сигнальных факторов будет воспринято растением как прогноз полного благополучия. Растение формируется изнеженным, и любая (даже непродолжительная) авария систем питания или кондиционирования грозит ему гибелью. Такая ситуация может возникнуть при выращивании в условиях высокой влажности воздуха или при круглосуточно неизменной температуре, поскольку сухой воздух и суточные колебания его температуры в естественных условиях являются сигналами заблаговременной подготовки к возможным невзгодам. Другой важный для практики эксперимента вывод состоит в возможности замены одного сигнального фактора другим. В известной степени взаимозаменяемы легкое (неповреждающее) воздействие пониженной влажности воздуха, снижение влажности корнеобитаемой среды, обогащение света ультрафиолетом и колебания температуры воздуха. Все они способствуют формированию более ксероморфного габитуса и повышению динамической устойчивости.
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Вы можете оставить свой комментарий авторизовавшись при помощи любой из представленный социальных сетей: