АгроСборник.Ру
С.Ф. Коваль, В.П. Шаманин "Растение в опыте"
При самых различных неблагоприятных воздействиях в растении наблюдается увеличение продукции этилена и абсцизовой кислоты (АБК), которую часто называю! гормоном стресса. Одновременно снижается количество ауксина, гиббереллинов и цитокинина. Такое изменение гормонального поля растения мы можем считать неспецифической реакцией тревоги организменного уровня.
Адаптация растительного организма имеет в своей основе статическую устойчивость отдельных клеток, но формируется она благодаря новообразованию структур (ростовые процессы) и обмену метаболитами внутри системы (транспорт). Благодаря этому, целый организм способен выживать в более тяжелых условиях, чем могут перенести его отдельные клетки или органы. На фоне умеренных воздействий поражение организма, определяемое по снижению продуктивности, бывает меньшим, чем установленное тестами повреждение изолированных тканей при этой же мере воздействия. Отличием динамической устойчивости от статической является относительно медленное развитие адаптационного процесса, благодаря чему максимальный эффект достигается через много дней после начала неблагоприятного воздействия. В силу этого химические анализы растения, проведенные через 5-10 дней после временного ухудшения экологических условий, отражают процессы внутриклеточной адаптации не в чистом виде, а с корректирующим влиянием организменной регуляции, т.е. не клеточную, а организменную реакцию. Формированию динамической устойчивости способствуют периодические ослабления нагрузки неблагоприятного фактора (отдых). Именно такой режим воздействия наблюдается в естественной обстановке, где дневная жара и дефицит влаги в воздухе сменяется ежесуточно ночным понижением температуры, а ночные заморозки -дневным потеплением. Аналогично: наиболее эффективны (в плане повышения устойчивости организменного уровня) не непрерывные режимы закалки, а те, в которых закаливающая нагрузка периодически сменяется благоприятными для роста условиями. В первое же время после неблагоприятного воздействия организменная устойчивость и скорость накопления биомассы бывает даже ниже, чем в контроле. Адаптация растения проявляется спустя некоторое время после репарации повреждений. В таблице 2.4 приведены данные по закалке прорастающих зерен пшеницы в непрерывном (повышение температуры от 30 до 70°С в течение 8 часов) - 1 режим; и периодическом режиме (ступенчатый нагрев от 30 до 70 °С с периодами оптимальной температуры) - 2 режим. Первый режим в большей степени выявляет клеточную адаптацию, а второй более благоприятен для формирования динамической устойчивости. К фазе кущения отмечено снижение темпов накопления сухого веса во втором варианте, т.к. адаптация на уровне организма еще не завершилась. Дополнительное тестирующее воздействие высокой температурой воздуха в эту фазу развития сильнее угнетало рост у контрольных растений. По этой причине различия между вариантами сразу после испытания отсутствуют. Через 10 дней после тестирующей нагрузки в полной мере реализовались эффекты организменного уровня, и вариант ступенчатой закалки вышел на первое место.
|
Таблица 2.4. Повышение жаростойкости пшеницы Лютесценс 758 при тепловой закалке семян [Игнатьев, 1973]
|
||||||||||||||||||||||||||
Проявлению динамической устойчивости способствует длительный, постепенно нарастающий, а не внезапный характер воздействия. Наибольшая адаптация организменного уровня наблюдается при режиме закаливания с чередованием периодов, неблагоприятных и близких к норме [Альтергот, 1981]. На этот момент следует обратить особое внимание, поскольку он связан с самой сущностью динамической устойчивости, формирующейся в процессе репарации полученных повреждений.
Выше отмечалось [Библь, 1965], что большинство факторов характеризуются наличием пространственного градиента по силе или по продолжительности действия. Благодаря этому, различные органы одного растения в естественной обстановке никогда не подвергаются одинаковым по силе внешним воздействиям. Этому способствует и различная устойчивость органов в связи с их возрастом и гистологическими особенностями. Так, в жаркий день в редком травостое у поверхности почвы температура бывает максимальной, а по мере удаления как в глубь почвы, так и в воздух она понижается. Иная картина наблюдается в случае ночного заморозка, когда охлаждение происходит за счет потери тепла земной поверхностью. В этом случае самая низкая температура отмечается у верхней границы травостоя. По мере углубления в травостой воздух становится все теплее, а подземные органы находятся в наиболее благоприятных температурных условиях.
Этому соответствует и картина повреждения. При постепенно нарастающей засухе в первую очередь завядают и отмирают нижние листья, а при заморозке - вершины побегов. Отмирание верхушек побегов и молодых листьев (при сохранении нижних) отмечается и при суховее, когда причиной гибели является интенсивное испарение, некомпенсированное поступлением воды по стеблю. Здесь мы имеем дело с градиентом водообеспечения в последовательном ряду органов, с перехватом воды теми листьями, которые расположены ближе других к корням. По этой же причине на фоне засухи в первую очередь засыхает верхушка листа, а основание пластинки, находясь в лучших условиях водоснабжения, дольше функционирует.
Из градиентного характера действия экологических факторов следует важный вывод о неравномерности поражения растения в любых природных условиях. В реальной обстановке почти всегда в пределах растения, наряду с поврежденными органами и тканями, имеются части с нормальным течением физиолого- биохимических процессов. Эти-то относительно благополучные в физиологическом отношении части и являются потребителями продуктов деструкции пораженных и отмирающих органов. Реутилизация продуктов частичного распада может происходи ть и на месте (в пораженных клетках), но значительная часть их транспортируется и вовлекается в обменные процессы слабо поврежденных, растущих органов. А это ставит динамическую устойчивость в зависимость от транспорта метаболитов внутри растения. Активность синтетических процессов в поврежденных органах понижена. Это способствует накоплению в них продуктов неглубокого распада (аминокислот, нуклеотидов). В отсутсгвии активного оттока репарация разрушенных структур хотя медленно, но произошла бы. Но когда скорость оттока превышает вовлечение метаболитов в репарационные процессы, происходит непрерывное опорожнение фонда продуктов распада (катаболитов). Равновесие распад-синтез сдвигается в направлении деструкции ультраструктур. В итоге, активно синтезирующие органы выкачивают вещества из стареющих или поврежденных [Альтергот, Махоткина, 1973] и тем самым ускоряют гибель последних. Это положение легко проверить, выполнив ряд анализов какой- либо формы свободных метаболитов в отсеченном органе и в гаком же органе, но находящемся в системе растения. У стареющего или поврежденного любым неблагоприятным фактором органа в отделенном от растения состоянии нарастает концентрация свободных метаболитов, поскольку транспорт их прекращается, а синтетические процессы не успевают использовать катаболиты. У растущего, физиологически активного органа в отсеченном состоянии концентрация метаболитов понижается (по сравнению с аналогичным органом в системе целостного растения), т.к. синтетические процессы преобладают.
Для эффективной реутилизации катаболитов в активных органах необходимы достаточно близкие к норме, обеспечивающие рост условия. Именно поэтому наибольшее повышение устойчивости достигается при многократном чередовании неблагоприятных и благоприятных условий [Альтергот, 1973]. Образующиеся в период активного роста органеллы, клетки, ткани и органы отражают в своей структуре предшествующее неблагоприятное воздействие и обладают повышенной статической устойчивостью. А это, в свою очередь, сказывается и на повышении устойчивости организма в целом.
Если условия для интенсивного роста отсутствуют, повышение устойчивости может быть достигнуто только за счет накопления и клетках осмотически активных веществ (в т.ч. и криопротекюрами), которые препятствуют потере воды поврежденными тканями и защищаю! белки от денатурации [Кушнер, 1969]. Важную роль может играть и накопление пула субстратов для дыхания. Кроме того, торможение раст яжения клсюк при неблагоприятных для роста условиях [Слейчер, 1970; Удовенко, Синельникова, 1972] приводит к образованию особо устойчивых мелкоклеточных тканей. Поражение обеспечивающих рост систем наступает раньше, чем снижение интенсивности фотосинтеза, а интенсивность дыхания еще дольше удерживается на стабильном уровне [Куперман, Хитрово, 1973]. Снижение расходов продуктов фотосинтеза на ростовые процессы приводит к переполнению ими клеток, что (наряду с накоплением катаболитов) обеспечивает повышение устойчивости.
Следует особо подчеркнуть, что высокая фотосинтетическая «мнимость и связанное с ней наличие пула метаболитов являются необходимыми факторами формирования динамической устойчивости. Общеизвестно, что для осенней закалки озимых культур требуется освещение достаточно высокой интенсивности. По нашим наблюдениям, степень торможения роста и накопления сухого веса у растений огурцов в результате 10-часового охлаждения до +4° С зависела от того, в какое н|Х!мя суток действовал неблагоприятный фактор (табл. 2.5). Мы объясняем уменьшение отрицательного эффекта при охлаждении в ночные часы наиичием в растениях достаточных запасов продуктов фотосинтеза, накопленных за прошедший день, тогда как при дневном охлаждении растения предварительно успевали расходовать в течение предшествующей ночи пул метаболитов на рост и дыхание.
|
Таблица 2.5. Динамика сухого веса (мг/раст) огурцов в зависимости от времени суток в момент охлаждения (+4° С х 10 ч.)
|
|||||||||||||||||||
Соответственно этому выбирается и методика закаливания для агротехнических целей или даже для получения здорового растения в эксперименте. Если до наступления неблагоприятного периода (каким может быть высадка рассады из парников, ослабление света с наступлением зимы и др.) осталось мало времени, следует применить торможение роста. Этот же прием используется в предвидении очень длительного неблагоприятного сезона. При заблаговременном закаливании или в случае перемежающегося действия неблагоприятного фактора рационально использовать стимуляцию роста.
На рисунке 2.1 дана предложенная В.Ф. Альтерготом [1969] физиологическая схема формирования динамической устойчивости, иллюстрирующая изложенные выше положения. Следует более подробно остановиться на блоке, помещенном в ее левом нижнем углу («Вторичные ростовые процессы»). Это понятие включает в себя новообразование органов с особенно сильной интенсивностью роста и стимуляцию регенерационных процессов. В некоторых случаях такое новообразование идет не из пазушных или апикальных почек, а из меристематических клеток сосудистых пучков. Такого рода «побеги израстания» томатов, возникающие после длительного периода жары из главной жилки листа обладают весьма высокой физиологической активностью, а растения из сформировавшихся на них семян имеют повышенную жароустойчивость.
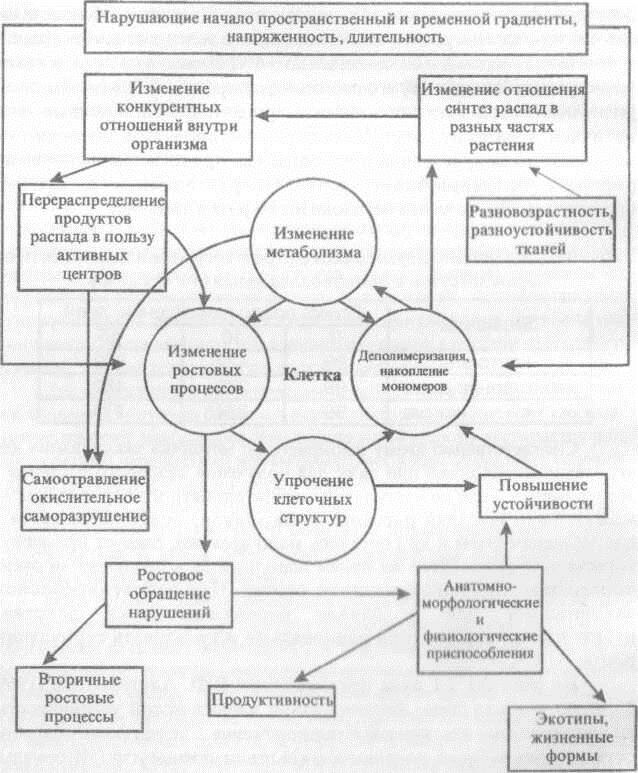
Р и с. 2.1. Схема саморегуляции адаптивных реакций
Используя это явление, можно стимулировать кориеобразование у стеблевых и листовых черенков различных видов растений. Указанный метод теплового повреждения верхушки черенка с целью стимуляции образования корней на базальном конце может с успехом использоваться для вегетативного размножения трудноукореняемых видов растений. Во всех случаях вторичного роста физиологической предпосылкой является частичная деструкция биополимеров в клетках некоторых органов при нормальной физиологической активности в других органах и клетках. Кроме ослабления конкуренции со стороны повреждаемого полюса и образования обильных пулов транспортируемых веществ, играет роль и совершенно уникальная физиологическая активность катаболитов, которая много выше, чем у любого входящего в катаболитный комплекс вещества, взятого в чистом виде. Здесь еще много неясного, но случаи бурной стимуляции роста при реутилизации растением продуктов неглубокого распада наводят на ряд размышлений. Не исключено, что в высокой физиологической активности катаболитов особую роль играет соотношение входящих в них компонентов (моно- и дисахаридов, аминокислот, нуклеотидов, ауксина и ряда витаминов). Другая сторона адаптации растения связана с выработкой такого мощного ингибитора, каким является абсцизовая кислота, в ответ на различные ухудшения условий существования. Можно предположить, что адаптивное значение этого гормона связано с торможением роста и последующим повышением устойчивости клеток, как и при обработке ретардантами.
Изложенные выше механизмы являются не более чем схемой этого сложного физиологического явления. Представления об организменном уровне адаптациии и устойчивости еще только формируются и действительная картина, безусловно, много сложнее наших нынешних представлений. В любых экспериментах с высшим растением необходимо учитывать возможности и границы динамической устойчивости растения. В заключение приведем фразу А. Поликара: «Самое опасное в этой области - это считать, что все обстоит просто».
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Вы можете оставить свой комментарий авторизовавшись при помощи любой из представленный социальных сетей: