АгроСборник.Ру

С.Ф. Коваль, В.П. Шаманин "Растение в опыте"
Продуктивность растения определяется взаимодействием физиологических процессов всех структурных уровней: клеточного, тканевого, органного, организменного. В применении к явлению продуктивности растения это означает невозможность свести его к простым закономерностям клеточной генетики или физиологии.
В качестве примера сошлемся на неудачные попытки ученых многих стран получить на основе донора Хайпроли высоколизиновые, ценные в кормовом отношении сорта ячменя. При этом исходили из стратегии переноса многими беккроссами гена высокого содержания лизина в адаптированные к местным условиям сорта. Считалось, что в процессе беккроссирования удастся получить сочетание свойственного Хайпроли высокого процента богатых лизином белков зерна с достаточной продуктивностью местных сортов. На языке генетики это обозначалось как разрыв вредной отрицательной корреляции признаков процента белка и массы 1000 зерен.
На деле оказалось, что повышенный процент высоколизиновых белков в зерне Хайпроли создается нарушением поступления метаболитов в растущие семена. В рассчете на одно зерно сокращается количество углеводов и, в меньшей степени, запасных низколизиновых белков. Как следствие, в щуплом зерне возрастает процент высоколизиновых конституционных белков. Таким образом, качество зерна Хайпроли является прямым следствием его щуплозерности и низкой зерновой продуктивности [Fijer, Fedak, 1984; Rybinski, Patyna. 1984; Stoy, Bertholdsson, 1984].
Другим примером ложной селекционной стратегии могут быть программы создания многолетней пшеницы (то же будет относиться и к другим однолетним культурам). Авторы сосредоточили внимание на получении трансгрессивных форм путем отдаленной гибридизации однолетних культур с многолетними растениями и видели главные препятствия достижению цели в проблемах цитогенетического плана. Но никто из них не задумался над тем, что зимовка растения требует расхода метаболитов на повышение осмотического давления в зимующих органах, на дыхание и репарацию повреждений в процессе зимовки.
Отток метаболитов в готовящиеся к зимовке вегетативные органы (вместо реутилизации их в зерно) неизбежно приведет к падению продуктивности. При постановке такой селекционной задачи прежде всего следовало решать вопрос о дополнительном источнике метаболитов для покрытия специфических потребностей в них многолетнего растения.
О неосуществимости данной селекционной легенды один из авторов говорит со знанием дела, поскольку он сам когда-то пытался получить многолетние томаты (размножаемые клубнями) в надежде освободить человечество от выращивания рассады.
Все попытки обнаружить «гены продуктивности» потерпели крах. Ныне признано, что наличие того или иного аллеля в генотипе еще не гарантирует ожидаемого вклада в продуктивность - он будет зависеть как от межгенных взаимодействий, так и от физиологической реализации генотипа в данных экологических условиях. И, напротив, отсутствие в генотипе желательного аллеля, конечно, исключает реализацию начинающегося с него физиологического эффекта, но не препятствует реализации продуктивности посредством иных физиолого-генетических путей.
На уровне организма большинство признаков детерминируется полигенно. Однако в любой регулирующей системе имеется ген, контролирующий лимитирующую реакцию. При этом фенотипически проявляются только эффекты аллельного состояния лимитирующего локуса. Остальные гены, сточки зрения внешнего наблюдателя, никак не проявляют себя в эксперименте. Принцип лимитирующего звена, сформулированный Ю. Либихом, может быть использован для описания регулировки сколь угодно сложных систем [Малецкий, 1982].
Приведенный ниже рисунок 3.4 изображает в общем виде некоторые возможные пути влияния генов (1,2,3,4) через их первичные продукты (5,6, 7,8) на физиологические процессы (9,10,11), а через них - на продуктивность (12) с учетом жологических условий произрастания (13). Гены (3, 4) определяю! физиологический процесс (9), который осуществляется при функционировании хотя бы одного из них, т.е. последний генетически стабилен. Вклад гена (2) в продуктивность через физиологический процесс (10) возможен только при воздействии на первичный продукт гена (6) физиологического процесса (9) и зависит от действия экологических условий
(13) на последний. Наконец, эффект гена (1) определяется как прямым вкладом физиологического процесса (11) в продуктивность, так и обоюдным влиянием процессов (10,11). Если экологические условия (13) будут оптимальными для данного генотипа, то реализуется потенциальная продуктивность. Любое отклонение климатических условий от оптимума приведет к снижению продуктивности. В этом случае мы говорим о реальной продуктивности растения в данных условиях. Если авторам удалось привести читателя в смущение запуганностью этой схемы, то они достигли поставленной ими цели, изображая сложность взаимодействия систем в растении.
Не следует забывать, что изображенные на схеме процессы, в свою очередь, являются реализацией иных систем генов. Таким образом, генетические системы, участвующие в формирвании урожая черезвычайно сложны и не поддаются анализу существующими сейчас методами.
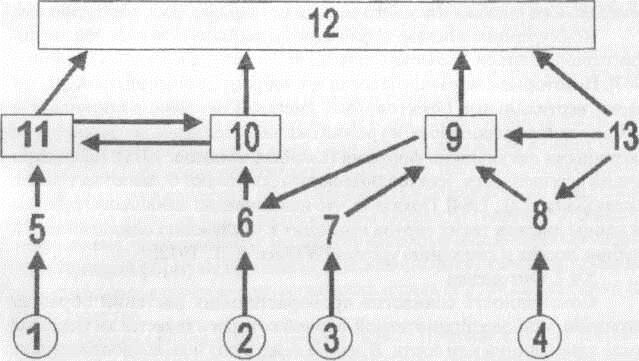
Рис 3.4. Схема сложных взаимодействий в организме (пояснения в тексте)
Читайте также
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()