АгроСборник.Ру

С.Ф. Коваль, В.П. Шаманин "Растение в опыте"
Основной ролью зеленого растения в биосфере является превращение неорганических веществ в органические с использованием энергии солнечного света [Мокроносов, 1983]. О космической роли зеленого растения в начале века писал еще К.А. Тимирязев. Биохимия и энергетика процесса фотосинтеза изложена в многочисленных монографиях, из которых сошлемся только на некоторые [Холл, Рас, 1983; Эдвардс, Уокер, 1986; Полевой, 1989; Шахов, 1993]. С математическими методами анализа продукционного процесса можно ознакомиться по книге И.В. Кармановой [1976]. Здесь мы рассмотрим фотосинтез только в той мере, насколько это необходимо для понимания механизмов формирования урожая.
Ассимиляция углекислого газа при фотосинтезе проходит в две фазы (Рис 3.1). В первой (световой) происходит поглощение кванта света хлорофиллом, диссоциация воды с образованием молекулярного кислорода и накопление энергии в АТФ и восстановленном НАДФ-Н (никотинамидадениндинуклеотид фосфат). Втемновой фазе (независящей от наличия света) за счет энергии АТФ и НАДФ-Н происходит присоединение молекулы углекислого газа к первичным акцепторам. Для ассимиляции одной молекулы С02 требуется не менее 8 квантов света, поглощенных пигментами. Энергия «моля квантов» фотонов (или энштейна) красного света (680 нм) равна 42 ккал, а 8 молей составят 336 ккал. Грамм-молекула глюкозы содержит 673 ккал, т.е. 112 ккал на грамм-молекулу ассимилированного С02. Следовательно, превращение энергии света в энергию органического вещества осуществляется с эффективностью 33 %. Для видимой части солнечного спектра при среднем значении энергии моля квантов около 50 ккал КПД ассимиляции С02 будет еще ниже. Если считать, что на долю фотосинтетически активной радиации (ФАР) приходится половина всего солнечного света, то эффективность фотосинтеза составит около 11 % [Эдвардс, Уокер, 1986]. Одним из источников потери энергии света в процессе фотосинтеза является слишком большая величина кванта. В видимом свете (720 - 380 нм) энергия одного энштейна кванта изменяется от 40 до 70 ккал при среднем ее значении 50 ккал. Интенсивность фотохимических процессов пропорциональна числу поглощенных фотонов, а не их энергии. Фотон в красной области спектра обладает меньшей энергией, чем фотон в синей части спектра, но оба они выполняют одинаковую фотохимическую работу. Если энергия поглощенного кванта слишком велика, то избыток ее рассеивается (переходит в форму тепла, излучается в виде флюоресценции). При поглощении кванта молекулы пигментов (хлорофиллы “а” и “Ь”, каротиноиды и др.) возбуждаются. Энергия возбужденного состояния может передаваться соседней молекуле хлорофилла, что приводит к дезактивации первой возбужденной молекулы и переходу в возбужденное состояние второй молекулы. Эффективность переноса энергии между молекулами хлорофилла близка к 100 %, тогда как от молекулы каротиноидов на хлорофилл “а” переносится около 40 % энергии возбуждения. Перенос осуществляется в направлении уменьшения энергии кванта: каротин (400 - 550 нм) -> хлорофилл b (650 нм) —> хлорофилл а (660 - 675 нм) ловушка (Р 680 или Р 700). Каротиноиды (как дополнительные пигменты) поглощают свет в синей и фиолетовой части спектра, недоступной для хлорофилла. Кроме того, эти пигменты защищают хлорофилл от окисления. Передача энергии от одной молекулы хлорофилла к другой может происходить сотни раз, пока она не будет принята особыми типами комплекса хлорофилла “а” с белком (Р 680 и Р 700), служащими ловушкой для квантов энергии. В реакционном центре содержится 200 - 400 молекул хлорофилла на одну молекулу ловушки.
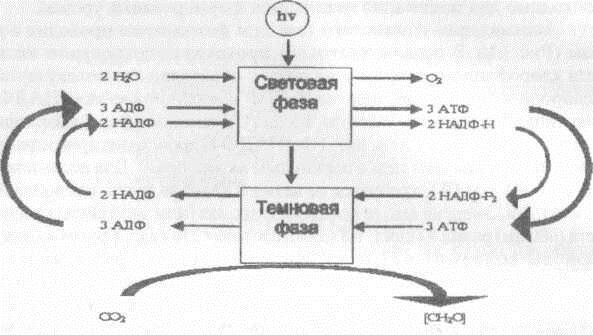
P и с. 3.1. Световая и темновая фазы ассимиляции СО фотосинтезе
Даже на прямом солнечном свету фотон поглощается молекулой хлорофилла не чаще одного раза за 0,1 сек. Таким образом, большую часть времени молекула хлорофилла «простаивает». Но при участии 200 собирающих кванты молекул хлорофилла ловушка будет возбуждаться каждые 0,0005 сек. Поэтому большинство молекул пигментов выполняют роль антенны, собирающей и передающей энергию света на небольшое количество хлорофилл-белковых комплексов Р 680 или Р 700, которые и осуществляют фотохимические реакции. Существуют разные пути восстановления С02 в фотосинтезе. Первый, известный как С-3 путь, состоит в присоединении С02 к рибулезо- 1,5-дифосфату (РДФ) с образованием двух молекул 3-фосфоглицериновой кислоты (С-3), которая после ряда преобразований дает начало моносахарам. На синтез одной молекулы глюкозы при этом затрачивается 12 НАДФ-Н и 18 АТФ. Этот механизм функционирует у большинства растений, для которых характерно наличие видимого фотодыхания, достаточно низкое световое насыщение фотосинтеза и неспособность ассимилировать С02 при низкой концентрации его в воздухе.
Одновременно с фотосинтезом в содержащих хлоропласты клетках идет и фотодыхание, т.е. светозависимое поглощение кислорода и выделение углекислого газа, которое не сопровождается окислительным фосфорилированием и не дает АТФ [Рейвн и др., 1990]. У С-3 растений фотодыхание усиливается при низком содержании С02 в воздухе и высоком парциальном давлении кислорода. Молекулы С02 и 02 конкурируют между собой за РДФ-карбоксилазу. При относительно высокой концентрации С02 в клетке преобладает карбоксиниронание РДФ, тогда как низкое отношение С02/02 благоприятствует окислению этого субстрата с образованием гликолевой кислоты. После ряда преобразований гликолевая кислота превращается в аминокислоту глицин, которая окисляется с образованием серина и освобождением СО2 В процессе фотодыхания рассходуется от 20-30 до 50 % первичных продуктов фотосинтеза, и на соответствующую величину уменьшается видимая интенсивность последнего. Конкуренция С02 и 02 за РДФ-карбоксилазу и связанное с этим фотодыхание приводят к наличию у С-3 растений углекислотного компенсационного пункта фотосинтеза (Рис. 3.2), ниже которого видимый баланс ассимиляции и диссимиляции С02 становится отрицательным.
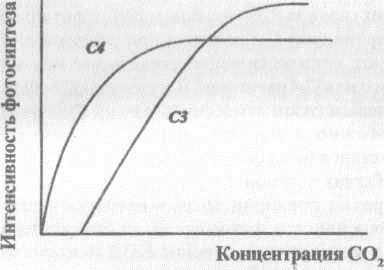
Рис. 3.2. Компенсационный пункт фотосинтеза у растений СЗ и С4
Иначе происходит восстановление С02 у растений с С-4 фотосинтезом, к которым относятся кукуруза, сорго, сахарный тростник, амарант, лебеда и некоторые другие. При этом молекула С02 присоединяется к фосфоенолпирувату (ФЕП) с образованием С-4 кислот. В таком виде ассимилированный углерод переносится из клеток мезофилла в хлоропласты клеток обкладки сосудистых пучков, где С02 вновь освобождается, а затем повторно ассимилируется в фотосинтезе по уже знакомому нам С-3 пути. Но при декарбоксилировании освобождается только половина того восстановительного потенциала, который необходим для ассимиляции одной молекулы С02. Таким образом, С-4 растения вынуждены расходовать на фиксацию одной молекулы С02 дополнительно две молекулы АТФ и, соответственно, больше квантов поглощенного света, чем растения С-3. Полагают, что С-4 путь создает накачку С02 и обеспечивает более высокую его концентрацию для конечного фотосинтеза по С-3 пути по сравнению с той, которая устанавливается диффузией его в лист из воздуха. В результате С02 перестает лимитировать фотосинтез, образование гликолевой кислоты и фотодыхание подавляются. Следствием является рост видимой интенсивности фотосинтеза по сравнению с растениями С-3, находящимися в одинаковых условиях освещения, а также целый ряд хозяйственных достоинств растений с С-4 путем фотосинтеза [Gifford, 1974; Monteith, 1978]. Указывают [Полевой, 1989], что С-4 растения могут при закрытых устьицах повторно использовать С02 фотодыхания. Закрытие устьичной щели в жаркие часы снижает испарение и существенно понижает транспирационный коэффициент растений С-4. Растения с С-4 путем фотосинтеза хорошо адаптированы к сильной инсоляции, повышенным температурам и к засухе [Рейвн, 1990]. Не исключено, что более распространенный фотосинтез С-3 возник в условиях атмосферы, богатой углекислотой и почти лишенной кислорода, в которой отсутствовала конкуренция этих газов за РДФ-карбоксилазу, и фотодыхание не имело места. Даже современные С-3 растения при содержании в воздухе 2 % кислорода имеют количественные характеристики интенсивности фотосинтеза, близкие к С-4 растениям. В современной богатой кислородом и бедной углекислым газом атмосфере древний С-3 путь недостаточно эффективен. Можно допустить, что мы являемся свидетелями эволюционного становления более эффективного С-4 пути фотосинтеза в различных семействах растений.
Неоднократно предпринимались попытки вести селекцию на повышение интенсивности фотосинтеза, но они не дали каких либо результатов. На молекулярном уровне КПД фотосинтеза ограничен квантовым выходом и особенностями спектров поглощения света пигментами.
Планы интенсификации карбоксилирования [Nasyrov, 1981] неосуществимы ввиду отсутствия изоферментного полиморфизма по ключевым энзимам фотосинтеза не только внутри видов (с чем и работают обычно селекционеры), но и в пределах высших растений вообще. Известные в литературе декларации о наличии генетического полиморфизма фотосинтеза оперируют только с мутациями, нарушающими нормальный процесс фотосинтеза и снижающими его продуктивность [Насыров, 1975], или же являются ошибками, связанными с тем, что автор [Насыров, 1981] принимает за генетический полиморфизм фотосинтеза межсортовое разнообразие по продуктивности, связанное с различной адаптацией генотипов к экологическим условиям. Надежды на увеличение нетто-фотосинтеза через депрессию темнового или фотодыхания не оправдываются, несмотря на наличие полиморфизма [Somerville, Ogren, 1982] по ряду локусов, связанных с превращением РДФ в процессе фотодыхания. Причина, видимо, в том, что в этих процессах происходит не только диссимиляция углерода, но и образование многих важных метаболитов. Равным образом, не известно ни одной удачной попытки перевести С-3 растения на С-4 путь фотосинтеза.
Известны успешные результаты отбора светолюбивых растений на теневыносливость [Кашевская, 1986] с целью создания генотипов, пригодных для произрастания в густых, оптически плотных посевах. Но достигнутый при этом результат зависит не от изменения физиологии фотосинтеза на клеточном уровне, он связан с процессами, происходящими на организменном уровне (К/хоз, потери на дыхание в нижних затененных листьях).
Читайте также
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()