АгроСборник.Ру

С.Ф. Коваль, В.П. Шаманин "Растение в опыте"
Совокупность совместно произрастающих растений образует фитоценоз, наиболее примитивной формой которого является загущенный посев одной линии или сорта. В ценозе действуют новые биологические законы, которые мы никаким образом не можем вывести из биологии клетки или отдельного растения.
Биологическое время ценоза протекает более медленно по сравнению сорганизменным временем. Поэтому с позиции ценотического уровня все составляющие его организмы статичны и постоянно находятся в режиме, наиболее оптимальном для данных экологических условий. С позиции же организма ценотические процессы протекают настолько медленно, что его изменения не заметны для более быстрых реакций нижележащих уровней.
Приведем несколько примеров влияния различных признаков растения на урожай ценоза.
Иммунитет к болезни повышает урожай в условиях эпифитотии, но не влияет на него вне заражения. Повреждение листьев болезнями приводит к ускорению старения их и более раннему созреванию (по сравнению с иммунным генотипом). При недостаточной продолжительности вегетационного периода иммунный генотип не успеет созреть, и вклад иммунитета в урожай станет отрицательным. В этом случае отсутствие устойчивости к болезни может дать тот же фенотипический эффект, что и гены, ускоряющие прохождение вегетации.
Сильное опушение листьев повышает устойчивость к грызущим вредителям, но волоски опушения хорошо удерживают капли дождя или росы, и этот дополнительный груз может вызвать полегание посева. Это можно наблюдать у яровой пшеницы Терция, которая отличается наличием густого опушения на листьях [Шаманин, 1994]. Сорт достаточно устойчив к полеганию, однако во влажные годы, под действием дополнительного груза удерживаемых капель дождя, в отдельных местах отмечается полегание посевов.
Ости у злаков облегчают отламывание чешуи колоса при механическом воздействии. На юге в сухую осень это способствует осыпанию зерна и снижает реальный урожай. На севере при сырой погоде остистые колосья лучше промолачиваются, и этот признак способствует уменьшению потерь зерна и тем повышает урожай.
Генетико-физиологические системы засухоустойчивости дают существенный вклад в урожай на фоне лимита влаги и не влияют на него при оптимальном водообеспечении. Напротив, все гены, увеличивающие число цветков и соцветий, могут увеличить урожай только при условии обильного питания или в разреженном посеве. В генетике это явление обозначено тяжеловесным термином «экологическое переопределение генетической формулы признака». Именно оно вынуждает нас проводить многолетние экологические испытания новых сортов до передачи их в производство.
Возникающие в фитоценозе новые признаки носят название групповых [Малецкий, 1982]. На отдельных растениях мы не можем оценить устойчивость посева к полеганию, процент погибающих в процессе вегетации растений, взаимное затенетие листьев в нижних ярусах травостоя, конкуренцию за влагу и минеральные элементы. На отдельно стоящем растении или на одно-двухрядковых делянках невозможно оценить и величину урожая в сомкнутом посеве. Подчеркиваем это особо, поскольку многие селекционера при отборе в селекционном питомнике ориентируются на “урожай” с однорядковой делянки.
Следует взять за правило: никогда не судить об урожае по однорядковым и даже по метровым делянкам. Крайние растения малых делянок (по сравнению со средними) находятся в несравнимо лучших условиях, т.к. используют влагу и плодородие межделяночных дорожек. Такое явление носит название краевого эффекта [Anslow, 1987; May et al., 1986]. Благодаря этому, растения крайних рядов часто бывают выше (по сравнению со средними), а поперечный профиль делянки имеет форму корыта. По наличию и глубине такого “корыта” можно визуально судить о неприспособленности сорта к экологическим условиям зоны, в которой ставит ся опыт.
В ценозе на долю одного растения приходится ограниченный объем воздуха и почвы. Этот объем вместе с приходящимися на долю каждого индивида ресурсами биотических и абиотических факторов жизни называется экологической нишей. Расширение своей экологической ниши за счет соседей мы определяем как внутривидовую и межвиловую конкуренцию. Генотипы обладают неодинаковой способностью захватывать объем воздушного или почвенного пространства, т.е. различаются по своей конкурентоспособности. Размеры экологической ниши уменьшаются по мере увеличения числа растений на единице площади или при снижении запасов экологических факторов. Соответственно этому, по мере загущения продуктивность падает.
Всходы и молодые растения в посеве с нормальным загущением еще не способны полностью использовать ресурсы своей экологической ниши. И этими избытками легко могут воспользоваться сорняки. По мере увеличения размеров растения оно все более полно осваивает свою экологическую нишу. Оптимальным будет такое загущения посева, при котором растения до конца вегетации полностью используют экологические ресурсы на формирование биомассы (урожая). Практически оптимальная густота посева выбирается по признаку полного освоения растениями наиболее лимитированного экологического фактора. В засушливых зонах таким будут запасы влаги, а во-влажных количество элементов минерального питания или (на фоне высоких доз удобрений) света, проникающего в глубину травостоя.
Изменение размеров экологической ниши в ходе онтогенеза является теоретической основой всех методов выращивания рассады. Молодые растения не в состоянии освоить всю площадь поля, которую они займут в дальнейшем. При загущенном размещении до высадки на постоянное место мы можем создать для них комфортные условия произрастания и защитить рассаду от неблагоприятных погодных условий. После высадки на постоянное место будет создана экологическая ниша, соответствующая размеру взрослого растения, а старт онтогенеза в благоприятных условиях теплицы, парника или хотя бы рассадника гарантирует успешное развитие до конца сезона. Высокая продуктивность отдельного растения еще не свидетельствует о большом урожае, который зависит от числа растений на единице площади и от ценотического взаимодействия, т. е. степени снижения продуктивности отдельных растений по мере загущения травостоя (Рис. 3.5).
Схемы опытов по разграничению эффектов конкуренции в зоне корней и в надземной части растений приведены в работе И.И. Симягина [1978], показавшего, что некоторые растения сильнее реагируют на сокращение воздушной экологической ниши, а другие - на сокращение почвенной. При этом не следует забывать, что в глубине травостоя не только уменьшается интенсивность светового потока, но и изменяется спектральный состав света [Тихомиров и др., 1991].
Переход от одиночного растения к более или менее загущенному посеву приводит к неизбежному падению средней продуктивности одного растения. Но до известного предела депрессия реальной продуктивности отстает от вызвавшего ее увеличения числа растений на единицу площади посева. В некоторых пределах урожай растет по мере загущения (Рис. 3.5). Это связано с тем, что занимаемая растением площадь используется неравномерно - периферическая часть меньше, чем центральная, и захват периферии другим растением снижает световое, водное и минеральное обеспечение слабее, чем это следовало бы из сокращения занимаемой площади.

Число растений на 1м2
Рис. 3.5. Изменение урожая в зависимости от плотности посева
Дальнейшее загущение посева выводит кривую урожая на плато, где проявляется взаимная компенсация между реальной продуктивностью и числом растений. Экологическая ниша при этом используется с максимальной полнотой, доступной данному генотипу. Загущение сверх компенсации приводит к быстрому падению урожая. Аналогично загущению посева действует и увеличение площади листьев на растении: возрастает затенение нижних ярусов и расход воды на транспирацию, т.е. крупное растение требует увеличения размеров экологической ниши.
Новейшие сорта и гибриды реализуют высокий урожай благодаря увеличенному числу растений не единице площади поля. С 1935 по 1876 года урожай зерна кукурузы возрос с 40 до 80 ц/га. Но эти гибриды имели преимущество по урожаю только при высокой плотности посева. Возникновение короткостебельных сортов зерновых культур обеспечило резкий рост урожая благодаря тому, что низкорослые сорта допускали более значительное загущение посева (без полегания и взаимного затенения) по сравнению с высокорослыми. Но создание агроценоза на основе интенсивных сортов приводит к изменению всех компонентов ценоза. Например, увеличивается биоразнообразие фитопатогенов [Монастырский, 1998].
Естественные фитоценозы включают растения многих видов, относящихся к различным жизненным формам. Сосуществование различных видов в ценозе возможно только благодаря ослабленной конкуренции между ними. Это имеет причиной как различные гребования к элементам питания, распространение корней на различной глубине в почве, так и неодинаковую форму надземных частей, что уменьшает взаимное затенение в густом травостое. Аналогичную картину мы наблюдаем и в искусственных смешанных посевах. С.Г. Вепрев с соавторами [1982] показали, что высокий урожай триплоидных популяций сахарной свеклы обеспечен не продуктивными свойствами триплоидных растений. Преимущества по урожаю таких популяций создаются различным габитусом листовых розеток ди- три- и гетраплоидных растений, что ослабляет световую конкуренцию в посеве (Рис. 3.6).
Конкурентные процессы в ценозе аналогичны по своим функциям распределению метаболитов между клетками в организме, т.е. составляют основу трофического регулирования системы. Но кроме распределения экологических ресурсов, имеется еще один регулятор в ценозе - это аллелопатические взаимодействия растений [Гродзинский и др., 1987]. Корневые выделения, эфирные масла, кутикулярные экскреты, вещества, вымываемые водой из живых листьев или мертвого отпада, фитоалексины, фитонциды и многие другие выделения способны подавлять жизнедеятельность некоторых компонентов ценоза и тем создавать благоприятную обстановку для других его составляющих. По своим функциям аллелопатичсские взаимодействия ценоза аналогичны гормональным регулирующим системам организма.
Достоинством смешанных посевов и естественных фитоценозов является и различная устойчивость их компанентов к неблагоприятным факторам. В примитивном сельском хозяйстве многие столетия практиковался смешанный посев озимой ржи с озимой пшеницей (суржа). В многоснежные зимы рожь выпревает сильнее пшеницы, а в малоснежные морозные годы она сохраняется лучше пшеницы. Таким образом, в любой год не происходит сплошной гибели смешанного посева. Выжившие растения за счет весеннего кущения частично используют экологические нишы погибших и снижение урожая от зимней гибели оказывается меньше ожидаемого.
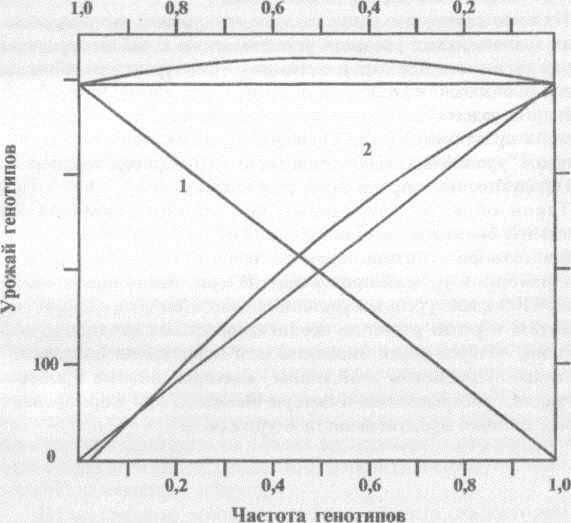
Рис. 3.6. Взаимодействие двух генотипов (1,2) в смешанном посеве, приводящем к повышению суммарного урожая
Генетическая гетерогенность многих селекционных сортов (см. главу 4) также является частным случаем смешанного посева. Володин с соавторами [1990] приводят несколько примеров гетерогенности сортов. Сорт озимого ячменя Нойга является смесью линий озимого типа и двуручек. Сорт яровой пшеницы Ремо состоит из двух линий, которые различаются даже внешне, но в смеси взаимно дополняют друг друга. Известны случаи, когда межсортовые смеси давали значительно более высокий урожай, чем чистые посевы любого из них. Но, к сожалению, в большинстве опубликованных опытов межсортовые смеси показывали не прибавку, а снижение урожая. Причина этого в нашем полном неведении принципов, по которым должны проводиться смешения сортов или линий. Неизвестно, следует смешивать сильного конкурента со слабым или же включать в смесь только слабых конкурентов. Не исключено, что решение этого вопроса может привести к радикальной смене направления селекционной работы - к переходу от селекции выравненных сортов к созданию синтетических смесей на основе ранее созданного материала.
Из всего сказанного выше следует, что урожай лимитируется в жестких экологических условиях устойчивостью к неблагоприятным факторам среды, а в близких к оптимуму - конкурентоспособностью растений. К сожалению, селекция на низкую конкурентоспособность специально не ведется ни в одном селекционном учереждении страны. Но повсеместно практикуется отбор в однорядковых делянках селекционных питомников “урожайных” кандидатов в сорта. Последнее, с точки зрения теории продукционного процесса, лишено смысла.
Таким образом, первичным источником формирования растительной биомассы является процесс фотосинтеза. Потенциальная интенсивность фотосинтеза огромна, но на пути реализации его в продуктивность и урожай происходят потери, связанные с низкой величиной Кхоз, конкуренцией растений в ценозе (посеве), торможением фотосинтеза и роста растения неблагоприятными экологическими факторами, потерей части биомассы при повреждени болезнями и вредителями. Предметом этой главы являлась система процессов образования, распределения и потери биомассы при формировании реальных величин продуктивности и урожая.
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()