АгроСборник.Ру

Физиолого-биохимические основы тактических приемов, используемых для защиты растений ячменя от pyrenophora teres sacc.
Полякова Н.В., к.б.н., Недведь Е.Л.
Институт экспериментальной ботаники ом. В.Ф. Купревича НАН Беларуси, г. Минск, Беларусь
Симптомы, возникающие при поражении листьев ячменя Р. teres и имеющие вид бурых пятен, получивших название сетчатой пятнистости, чрезвычайно вредоносны. В отдельные годы потеря урожая от сетчатого гельминтоспориоза составляет до 20% и более [1].
Оптимизировать формирование урожая за счет снижения функциональной активности возбудителя сетчатого гельминтоспориоза или за счет устранения его из фитопатосистемы невозможно без знания метаболических процессов, изменяющих под влиянием инфекции интенсивность их протекания. Сочетание этих процессов с условиями выращивания посевов ячменя рассматривается нами как основополагающий принцип при оценке степени соответствия новой среды существования патогена его генетической природе и биологическим особенностям, а также при разработке рациональных противоинфекционных средств фитозащиты. В таком аспекте исследования взаимоотношений, складывающихся в системе хозяин - патоген, практически не проводились.
Наше внимание было сосредоточено на изучении роли фитогормонов в формировании тканями ячменя ответной реакции после взаимодействия их с P. teres. Этот подход был выбран нами в соответствии с атрибутами, характерными для регуляторов роста и развития, а конкретно с их способностью активировать определенные группы ферментов, ускорять транспорт метаболитов и осуществлять их переориентацию к месту накопления [2].
Инфицирование ячменя Р. teres сопровождается в местах внедрения патогена изменениями гормонального баланса [3]. Сам патоген способен выделять в среду роста широкий спектр гормональных соединений [4].
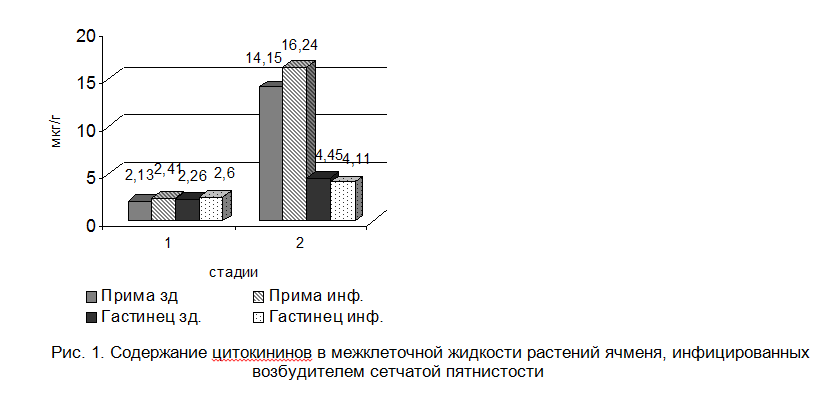
В межклеточной жидкости тканей ячменя устойчивого сорта происходит повышение уровня цитокининов, тогда как у восприимчивого сорта - его снижение (рис.1). Поскольку P. teres продуцирует цитокинины, среди которых преобладает зеатинрибозид, есть все основания предполагать, что основной вклад в наблюдаемое эндогенное повышение концентрации цитокининов вносит гриб. Видимо, зеатинрибозид является одним из главных факторов внедрения и проникновения гриба в ткани устойчивого сорта ячменя. Известно, что экзогенные цитокинины способны подавлять активность хитиназы - фермента, защищающего от инфекции растения [5], также продуцирование активных форм кислорода, индуцирующих сверхчувствительный некроз растений, отторгающий патоген [6]. Однако наши исследования показали, что увеличение содержания активных форм кислорода в ответ на инфекцию P. teres тканей ячменя не предотвращает, а напротив, стимулирует инфекционный процесс. Более того, цитокинины участвуют в трансдукции сигнала при различных стрессах, в том числе при внедрении патогена. Увеличение уровня цитокининов связано и с некоторыми морфогенетическими изменениями растений [7]. Наиболее вероятным объяснением данного явления нам представляется положение о том, что происходящее за счет грибного гормона возрастание уровня цитокининов благоприятствует продвижению P. teres по межклетникам листа и израстанию грибницы, с помощью которой обеспечивает наилучший доступ к питательным веществам.
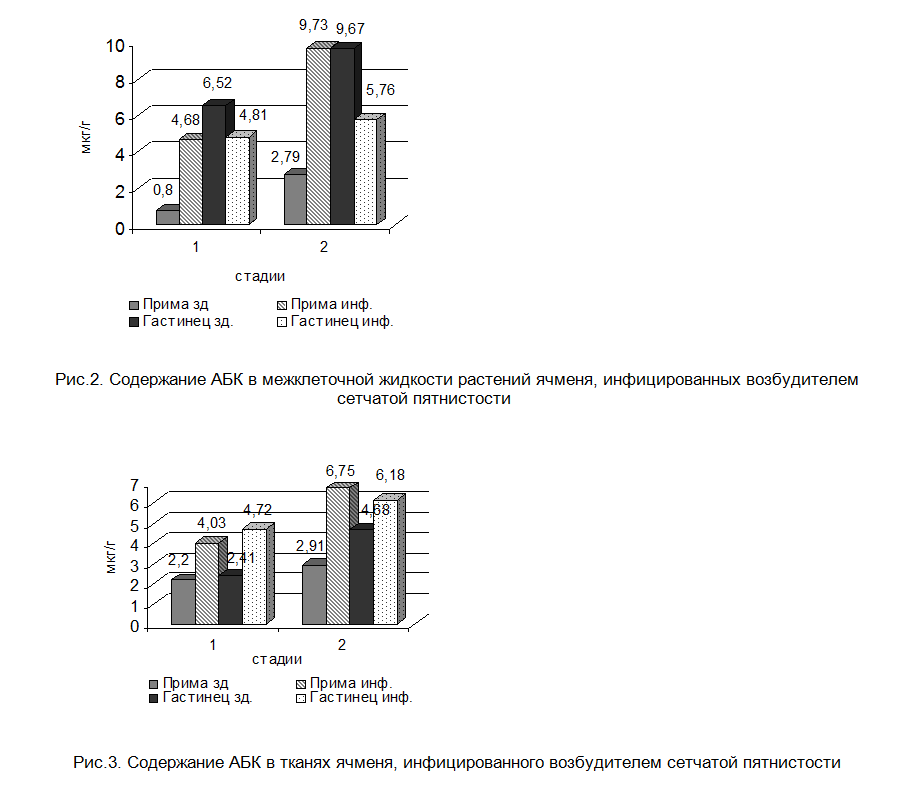
Инфицирование ячменя P. teres в наших опытах вызывало также резкое повышение уровня АБК в устойчивом сорте на всех стадиях роста. В восприимчивом генотипе ячменя тенденция к повышению содержания АБК значительно снижена и по мере развития симптомов при переходе к следующей стадии. Иная картина характерна при определении содержания АБК в межклеточном пространстве. В апопласте устойчивого сорта ячменя отмечалось резкое многократное увеличение уровня этого гормона, тогда как у аналога восприимчивого сорта - его падение (рис.2, 3).
Процесс узнавания патогена, возникающий в месте внедрения его в ткани хозяина инициирует активность системы сигнализации, в ходе функционирования которой во взаимоотношения организмов включается комплекс защитных реакций на локальном и системном уровнях. В составе последних изменения метаболизма инфицированного растения и, в частности, гормональной и окислительно-восстановительной составляющих его. Нами предполагается, что взаимосвязь между соотношением окислительные/антиокислительные процессы и балансом ростостимулирующая и ростоингибирующая гормональная активность играют значительную роль в формировании фитопатосистемы ячмень - Pyrenophora teres Sacc.
Согласно литературным данным, инфицирование растений грибными патогенами приводит к сдвигу равновесия между оксидантами и антиоксидантами в сторону усиления свободнорадикальных процессов [8, 9]. Это же происходит и в исследуемой нами системе ячмень - возбудитель сетчатой пятнистости, где между партнерами устанавливаются факультативные паразитические связи. В результате запускается интенсивное некрозообразование клеток растения, но оно, в отличие от облигатных патосистем, создает благоприятную среду обитания для P. tеres - некротрофа по способу питания. Устойчивость же проявляется в период пребывания ткани хозяина в жизнедеятельном состоянии. От степени пролонгированности данного состояния зависит продолжительность стадии -биотрофности P. teres, а следовательно, срок функционирования болезнеустойчивости.
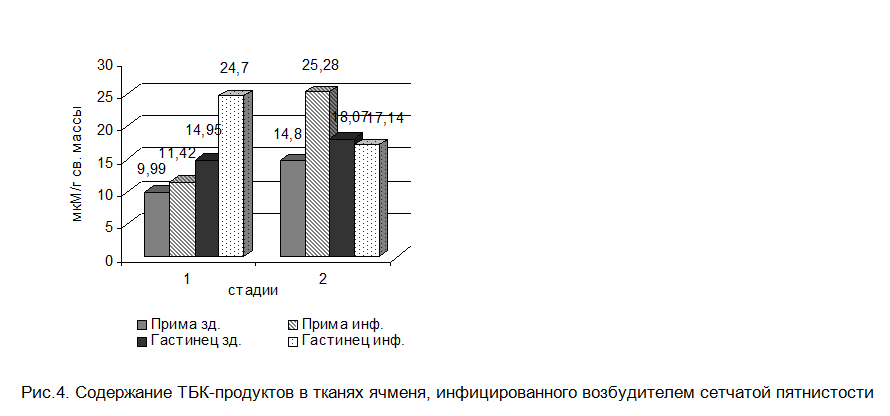
Эти данные согласуются и с характером изменений в гормональной активности растения-хозяина, При формировании факультативных отношений наблюдается прямая зависимость между повышением уровня АБК и интенсивностью перекисного окисления липидов мембран (рис.4). АБК, как сигнальная молекула, сдвигает гормональный баланс в сторону снижения ростостимулирующей активности и разрушения мембран растительных клеток. По-видимому, увеличение содержания АБК в инфицированных P. teres листьях ячменя приводит к сдвигу метаболизма растения-хозяина в сторону усиления окислительных процессов, к некрозообразованию и развитию болезни. Долговременное поддержание высокой концентрации АБК сопровождается формированием совместимых взаимоотношений между патогеном и хозяином (даже у устойчивого сорта) и обусловлено значительным продуцированием данного гормона самим грибом. Вероятно, роль грибной АБК заключается в том, что она служит сигналом, трансдукция которого ингибирует рост мицелия, а следовательно, переход гриба к спороношению и способствует многократному воспроизводству его в течение периода вегетации.
Итак, физиолого-биохимическую основу тактики, используемой для защиты посевов ячменя от пятнистостей листьев, составляет определенное сочетание гормонального и редоксстатуса растения, складывающееся при поражении листьев P. teres. Усиленная скорость генерации активных форм кислорода напрямую контролируется уровнем АБК. Менее эффективен в этом процессе уровень цитокининов. Оба механизма приспособлены, наряду с защитой растений от патогенов, к реализации и других жизненных функций: узнавание чужеродных метаболитов и их детоксикация, индуцирование генов, включающих в процесс межвидовой интеграции проявление более поздних защитных реакций в клетках растений, взаимное регулирование синтеза свободнррадикальных и гормональных соединений.
Литература
1. Гайке М.В., Бенкен А.А. Сетчатый гельминтоспориоз ячменя // Сельхозяйственная биология. -1977. – Т 6/2 . - С. 193-198.
2. Кулаева О.Н. Идеи А.Л. Курсанова в исследовании фитогормонов // Физиология растений. - 2002. - Т,49. №1. - С. 85 – 86.
3. Серова З.Я., Волынец А.П., Полякова Н.В. Гормональные соединения в межвидовых отношениях ячменя и возбудителя сетчатого гельминтоспориоза // I Всерос. конф. по иммунитету растений к болезням и вредителям: Сборник научных докладов. - С-Пб, 2002. - С. 49-50.
4. Полякова Н.В. Физиологически активные вещества фитопатогенного гриба Helminthosporium teres Sacc: Автореферат дис. ... канд. биол. н., - Минск, 1993. - 21с.
5. Sihishi H., Mohnen D., Meins F. Regulation of the Response of Phaseolus vulgaris to a Fungal Elicitor // Plant Cell Physiol. - 1990. - V. 31. - P. 687-675.
6. Мерзляк М.Н., Жиров В.К. Свободнорадикальное окисление в хлоропластах при старении растений // Итоги науки и техники. Сер. Биофизика. - М.: ВИНИТИ, 1990. - Т. 40. - С. 101 –135.
7. Максимов И. В., Ганиев P.M., Хайруллин P.M. Изменение уровня ИУК, АБК и цитокининов в инфицированных Tilletia caries проростках пшеницы // Физиология растений. - 2002. – Т. 49. №2. - C. 248-252.
8. Mehdy M.C. Active oxygen species in plant defense against pathogens // Plant. Physiol. - 1994. - V. 105. - P. 467-472.
9. Wojtaszek P. Oxidative burst: an early plant response to pathogen infection // Biochem. J. -1997. – V. 322. - P. 681-692.
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()