АгроСборник.Ру

Создание секалотритикум - экспериментальный ароморфоз и эффективный путь расширения генофонда амфидиплоидов пшеницы с рожью
Белько Н.Б., к.б.н., Гордей И.А., д.б.н., Хохлова С.А., к.б.н., Щетько И.С., Быченко А.П.
Институт генетики и цитологии НАН Беларуси, г. Минск, Беларусь
Селекционно-генетический анализ созданного генофонда тритикале показал, что у тритикале недостаточно реализован генетический потенциал адаптивности ржи. Тритикале, как правило, уступают озимой ржи по морозо- и зимостойкости, устойчивости к грибным болезням и более требовательны к плодородию почв. Урожайность тритикале варьирует в зависимости от зоны и года возделывания, что обусловлено недостаточной экологической пластичностью и слабой устойчивостью растений к биотическим и абиотическим факторам. Причиной недостаточной экологической пластичности используемых в сельскохозяйственном производстве пшенично-ржаных амфидиплоидов является ингибирование в гибридном ядре генетических систем ржи, определяющих высокую адаптивность растений, вследствие взаимодействия их с количественно преобладающими геномами и цитоплазмой пшеницы.
Дальнейший прогресс в селекции этой культуры может быть связан с решением проблемы усиления экспрессии ржаного генома в полигеноме амфидиплоидов ржи с пшеницей, расширением и обогащением генофонда исходного материала на основе вовлечения видового потенциала пшеницы и ржи путём синтеза амфидиплоидов различного геномного состава и ядерно-цитоплазматической структуры с последующей интрогрессивной и рекомбинационной селекцией. В этой связи особый интерес представляет создание рекомбинантных форм тритикале с цитоплазмой ржи [1,2] и их хромосомно-замещенных линий. Синтез таких форм будет способствовать оптимизации цитоплазматического фона для экспрессии геномов исходных видов в полигеноме гибридов и расширению генетического разнообразия исходного материала вследствие эффектов замещений хромосом и новых ядерно-цитоплазматических взаимодействий.
В данной работе представлены результаты цитогенетического анализа гибридов F1 (тетраплоидная рожь, 4х=28 х гексаплоидное тритикале, 6х=42), идентификации кариотипов методом дифференциального окрашивания хромосом созданных секалотритикум и их замещенных форм в связи с морфотипом и продуктивностью растений.
В качестве исходного материала использованы районированные и перспективные сорта тетраплоидной озимой ржи (RRRR, 4x=28), формы гексаплоидного тритикале (AABBRR, 6x=42) и сорта мягкой озимой пшеницы (AABBDD, 6x=42). Создание секалотритикум осуществлялось по разработанному нами методу, основанному на использовании в скрещиваниях с рожью гексаплоидных тритикале в качестве источника геномов пшеницы [2-4]. Хромосомно-замещенные линии секалотритикум получали путём интрогрессивной гибридизации секалотритикум с мягкой пшеницей. Исследование микроспорогенеза у гибридов проводили на окрашенных ацетокармином давленых препаратах. Хромосомно-замещенные линии секалотритикум идентифицировали с помощью метода дифференциального окрашивания хромосом.
Установлено, что определяющим этапом формирования генома секалотритикум являются ржано-тритикальные пентаплоиды F1 (RRABR, 5х=35). Мейоз у них характеризуется рядом специфических особенностей вследствие специфики геномного состава – один диплоидный (RR) и три гаплоидных (ABR) генома - и взаимодействия генетических систем контроля микроспорогенеза ржи и пшеницы. Основные из них: 1) высокий уровень гомеологичной конъюгации хромосом (до 15 бивалентов на клетку вместо 7), связанный с ингибированием Ph-системы пшеницы бивалентной конъюгации гомологов в условиях ржаной цитоплазмы и трёх геномов ржи и уменьшением дозы Ph-гена; 2) эквационное деление части унивалентов (до 7) в А1 вследствие перехода от однополярной ориентации хромосом к диполярной или нарушения связи одной из хроматид с полюсом под действием тройной дозы P/Edu генов; 3) неравномерное распределение генетического материала по клеткам, образование диад и тетрад (от 72,2 до 90,9%) с микроядрами.
Специфические особенности мейотического деления, вызванные межгеномными и ядерно-цитоплазматическими взаимодействиями, координируют мейоз унивалентных хромосом, делая его в целом более регулярным, чем у истинных гаплоидов, и приводят к внутри- и межгеномным рекомбинациям, неравномерному расхождению и интенсивной элиминации генетического материала. Это обусловливает формирование у ржано-тритикальных гибридов F1 относительно высокого процента жизнеспособных гамет и зерновок (фертильность пыльцы от 5,1 до 13,0%, фертильность колоса при свободном опылении - в среднем 4,9 зерна на колос) с различным числом и геномным составом хромосом. В результате в F1BC1 и последующих поколениях обеспечивается получение ржано-пшеничных амфиплоидов в достаточном объёме с широким спектром изменчивости (табл. 1).
Таблица 1 Распределение ржано-тритикальных амфиплоидов F1BC1 по числу хромосом
|
Число хромосом |
35 |
36 |
37 |
38 |
39 |
40 |
41 |
42 |
43 |
47 |
Растений |
|
Частота,% |
14,2 |
4,2 |
17,4 |
22,7 |
12,0 |
14,7 |
2,5 |
11,7 |
0,1 |
0,4 |
674 |
У гибридов F1BC1 число хромосом варьировало в пределах 33-47. Большая часть растений содержала 35, 37, 38, 39, 40 и 42 хромосомы. Большинство 42-хромосомных растений имели несбалансированный кариотип по наборам хромосом пшеницы и ржи, характеризовались низкой фертильностью. Сбалансированные по наборам хромосом, цитологически стабильные и фертильные гексаплоиды составляли около 1%.
На основе 42-хромосомных растений, выщепившихся в F2BC1 и F1BC2 в скрещиваниях 8 сортов тетраплоидной озимой ржи (Радзіма, Новосибирская, Верасень, Пуховчанка, Ясельда, Ислочь, Калинка, Fernando) и 10 форм гексаплоидных тритикале с использованием разработанного в лаборатории метода создан генофонд ржано-пшеничных амфидиплоидов с цитоплазмой ржи, включающий более 200 семей F1-F4 секалотритикум. Установлено, что степень проявления морфологических, хозяйственно-ценных признаков и свойств у ранних поколений созданных секалотритикум варьировала в зависимости от комбинации скрещивания, а также от семьи и конкретного растения в пределах одного и того же гибрида. Например, у линии секалотритикум 109-4, полученной на основе сорта ржи Новосибирская и тритикале Л 246, масса 1000 зерен составила 43,1, а у линии этой же комбинации 110-2 – 47,0 г, т.е оказалась выше на 9,3%. В то же время для линии 110-2 характерны в среднем более низкие озерненность и масса зерна с растения. Линя 109-1 созревала на неделю позже линий 109-4 и 110-2. Сравнение коэффициентов вариации одноименных признаков позволило установить большую степень изменчивости элементов структуры урожая у созданных секалотритикум по сравнению с тритикале. При этом максимальные варианты у секалотритикум имели более высокие значения признаков, чем максимальные варианты у тритикале. Выявленные между линиями и растениями одного и того же гибрида различия в степени проявления признаков и свойств свидетельствуют о наличии генотипической изменчивости у ранних поколений созданных ржано-пшеничных амфидиплоидов, которая обусловлена рядом факторов. Из основных факторов, кроме сложного геномного состава ржано-тритикальных амфиплоидов F1 (RRABR, 5x=35) и F1BC1 и специфичности протекания у них мейоза, следует выделить модификации хромосом, возникающие у гибридов отдаленных скрещиваний и разнообразие аллелей геномов A, B и R исходных сортов тритикале и ржи, используемых при получении секалотритикум. В результатье этих процессов у ржано-тритикальных амфиплоидов F1, F1BC1, F2BC1 и F1BC2 в процессе мейоза образуется большое разнообразие гамет, объединение которых при оплодотворении приводит к разнообразию зигот, гетерогенности последующих поколений и сложному характеру расщепления. Высокая гетерогенность ранних поколений секалотритикум и широкий спектр изменчивости признаков продуктивности являются базой для создания генетического разнообразия исходного материала секалотритикум, отбора генотипов с комплексом элементов продуктивности и создания высокоурожайных и экологически пластичных линий.
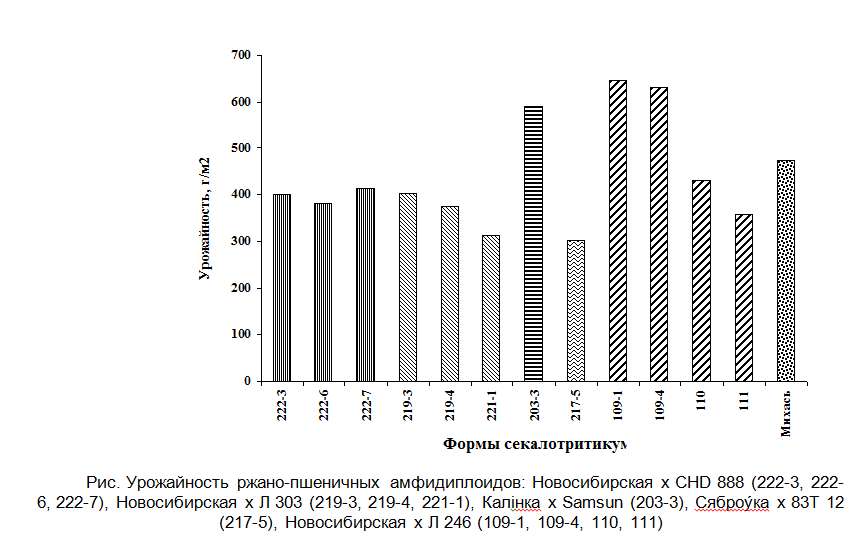
Выделено 24 цитологически стабильных форм секалотритикум. С использованием метода дифференциального окрашивания хромосом проведен анализ кариотипов некоторых ржано-пшеничных амфидиплоидов (табл. 2). Установлено, что исследованные секалотритикум являются геномно-сбалансированными гексаплоидными формами. Их кариотипы представлены полными наборами хромосом A, B геномов пшеницы и R генома ржи. Выявлены характерные особенности в рисунках дифференциального окрашивания хромосом ржи, затрагивающие как терминальные, так и интеркалярные C-блоки, которые сформировались вследствие межгеномных и ядерно-цитоплазматических взаимодействий и специфичности R-геномов, привнесённых в полигеном секалотритикум исходными тритикале и рожью. Некоторые секалотритикум характеризовались более высокой урожайностью по сравнению со стандартом – сортом тритикале Михась, которая обеспечивалась более высокой жизнеспособностью растений и высокими средними значениями элементов структуры урожая (рис).
С целью расширения генетического разнообразия и улучшения хлебопекарных качеств секалотритикум проводили реконструкцию их генома. Предполагалось, что интрогрессия в кариотип секалотритикум хромосом D-генома пшеницы позволит сохранить признаки, присущие ржаному геному: адаптивность, зимостойкость, устойчивость к болезням, и одновременно улучшить хлебопекарные свойства за счёт качества клейковины и белкового состава зерна.
Таблица 2 Хромосомный состав кариотипов и морфо-биологические особенности секалотритикум и хромосомно-замещенных линий
|
Линия |
Происхождение |
Хромосом, в том числе по геномам |
Идентификация замещенных хромосом |
Морфотип линии |
Биологические особенности |
Продуктив-ность, г/м2 |
||||||||
|
общее |
пшеницы |
ржи |
||||||||||||
|
A |
B |
D |
R |
|||||||||||
|
Секалотритикум |
||||||||||||||
|
109-1 |
Новосибирская х Л.246 |
42 |
14 |
14 |
- |
14 |
|
Высота 115см. Колос длинный и рыхлый |
Позднеспелая, устойчивая к мучнистой росе и септориозу |
646,4 |
||||
|
203-3 |
Калинка х Самсун |
42 |
14 |
14 |
- |
14 |
|
Высота 118 см. Колос плотный |
Среднеспелая, озерненность колоса высокая |
590,0 |
||||
|
Хромосомно-замещенные линии |
||||||||||||||
|
220 |
(Верасень х Л374) х Л160 х Л160 |
42 |
14 |
14 |
14 |
- |
Полное замещение ржаного генома – D/R |
Характеризуется низкой зимостойкостью |
Аллопшеница. Поражается мучнистой росой и септориозом |
268,8 |
||||
|
227 |
(Новосибирская х Л246) х Гармония х Гармония |
42 |
14 |
14 |
12 |
2 |
Пара хромосом 2R, остальные замещены на хромосомы D-генома |
Пшеничного морфотипа, фертильность до 90%. Линия среднеспелая |
Линия устойчивая к септориозу |
401,6 |
||||
|
129 |
(Папараць х AD-60) х Луцковлянка х Луцковлянка |
42 |
14 |
14 |
2 |
12 |
Замещение 1D/1R |
Морфотип секало- тритикум, средне-спелая, высокофер-тильная (95%) |
Линия устойчивая к грибным заболеваниям |
462,9 |
||||
|
217 |
(Новосибир-ская х Л246) х Копылянка х Копылянка |
42 |
14 |
14 |
|
14 |
Присутствуют 14 хромосом ржаного генома, имеются транслокации |
Морфотип секало-тритикум. Растения выровненные, фертильные |
Линия устойчивая к септориозу |
707,7 |
||||
|
Тритикале Михась |
42 |
14 |
14 |
|
14 |
|
|
|
472,9 |
|||||
В результате интрогрессивной гибридизации секалотритикум с мягкой озимой пшеницей получена серия хромосомно-замещённых форм ржано-пшеничных амфидиплоидов на цитоплазме ржи с различным сочетанием хромосом R- и D-геномов и степенью проявления хозяйственно-ценных признаков и свойств (табл. 2). Установлено, что количество замещенных хромосом в кариотипе линий варьировало от 1 до 7 пар ржаного генома на хромосомы D генома пшеницы. Кариотипический состав интрогрессивных линий определял морфотип растений: замещения 1 и 2 пар хромосом ржаного генома обуславливает формирование растений с морфотипом исходных секалотритикум, а замещение 5-7 пар ржаных хромосом – с морфотипом пшеницы. Выделены интрогрессивные линии секалотритикум с различным сочетанием хромосом
R- и D- геномов (линия 129 – замещена 1R хромосома, морфотип секалотритикум; линия 227 – замещена 2R хромосома, морфотип секалотритикум; линия 220 – полное замещение ржаного генома на хромосомы D-генома пшеницы, морфотип пшеницы). Практическое значение имеют линии с замещением не более двух пар ржаных хромосом на хромосомы D-генома пшеницы. Замещения двух пар хромосом ржи не снижает продуктивность, адаптивность и устойчивость растений к болезням.
Проведенные исследования подтверждают целесообразность создания ржано-пшеничных амфидиплоидов с цитоплазмой ржи (секалотритикум) в связи с повышением адаптивности, расширением и обогащением генофонда амфидиплоидов пшеницы с рожью и высокую эффективность разработанного метода их синтеза. Полученные результаты важны для выяснения роли и эффектов взаимодействия генетических систем контроля процесса микроспорогенеза и отдельных хромосом исходных видов у отдаленных гибридов. Созданный генофонд секалотритикум и хромосомно-замещенные линии представляют интерес для практической селекции и генетических исследований.
Литература
1. Гончаров С.В., Туровский А.И. Использование тетраплоидной ржи для получения рекомбинантных форм тритикале // Новое в селекции и семеноводстве сельскохозяйственных культур. - 1987. - С. 152-155
2. Гордей Г.М., Гордей И.А., Новикова Л.В., Клименко Е.П. Способ получения секалотритикум. А. С. №1734602 А1. - 1992.
3. Гордей И.А., Гордей Г.М., Новикова Л.В. Генетические основы создания тритикале (хТритикале): Создание ржано-пшеничных амфидиплоидов (секалотритикум) // Генетика. – 1996. - Т.32. С. 783-787.
4. I.A.Gordei, N.B.Belko, S.A. Khokhlova Cytogenetic analysis of genome formation in rye-wheat amphploids // 11th International Congress on Genes, Gene Families and Isozymes, june 30-july 4, 2001, Stocholm, Sweden. - P.58.
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()