АгроСборник.Ру

Гетерозис и коэффициент доминирования по признаку массы гибридных семян (F1) картофеля
Гетерозис (от греческого слова «хетероиосис» — «изменение», «превращение») — это ускорение роста, увеличение размеров, повышение жизнеспособности и продуктивности гибридов первого поколения по сравнению с родительскими организмами. При скрещивании организмов с разной наследственностью, происходит биохимическое обогащение гибрида, у него усиливается обмен веществ, что и вызывает проявление гетерозиса. В последующих поколениях эффект гетерозиса постепенно затухает. У вегетативно размножаемых растений (картофель, плодовые и ягодные культуры) возможно закрепление гетерозиса в потомстве (Ермишин А.П., 1998; Симаков Е.А., 2010; Шанина Е.П., 2012).
По сообщениям З.В. Абрамова (1985), гетерозис – явление более мощного развития гибридов первого поколения по сравнению с родительскими формами – линиями и сортами. Термин гетерозис был предложен Шеллом в 1914 г., но явление более мощного развития гибридов по сравнению с родительскими формами у многих видов растений, было описано Дарвином в 1862 г. (цитат. по Абрамовой, 1985).
А. Густафссон (Gustafson А., 1946) описывает три типа гетерозиса у растений:
1. репродуктивный (цветков и соцветий),
2. соматический (стеблей, побегов, листьев, клубней, корнеплодов) и
3. адаптивный (зимостойкости, засухоустойчивости, скороспелости).
Многие авторы сообщают (Будин К.З., 1986; Камераз А.Я., 1973; Яшина И.М., и др., 1973; Kaushik S.K. et al.,1996; Gupta K.S.et al., 2004; Luthra S.K.et al., 2006), что далеко не все исчерпаны возможности совершенствования традиционных методов селекции при гетерозисе, связанных, в частности, с выделением лучших родительских форм, их гибридизацией, отбором гетерозисных гибридов в ранних поколениях.
Как сообщает Х. Росс (1989), гетерозис у картофеля проявляется в тех случаях, когда потомство превосходит лучшего родителя или среднюю для обоих родителей.
Использование гетерозиса – наиболее важная цель практической селекции картофеля. Наследование гетерозиса обусловлено малыми генами или побочными эффектами главных генов. Их совместное действие может проявляться аддитивным (ОКС) или неаддитивным (СКС) способом. В большинстве случаев действуют оба фактора. Установлены высокие значения ОКС и СКС по общей урожайности и ее двум компонентам - числу клубней на куст и средней массе одного клубня, взаимодействие которых мультипликативно.
Гетерозис у картофеля в основном базируется на неаддитивном взаимодействием генов. Он включает как внутрилокусное (сверхдоминирование), так и межлокусное (эпистаз) взаимодействие генов и аллелей. Таким образом, выщепление гетерозисных сеянцев в популяции бывает, по – видимому, наибольшим, при трех условиях:
1- предельно низком коэффициенте инбридига у родительских форм;
2- максимальном количестве локусов, различающихся по аллелям;
3- различном генофонде родительских форм, что увеличивает аллельное разнообразие (широкая гибридизация).
Решение проблемы повышения эффективности селекции картофеля, видится в сочетании традиционных и новых генетических подходов. Успех селекции в наибольшей степени зависит от разнообразия исходного материала, с хозяйственно-полезными признаками и другими биологическими свойствами. В связи с этим, нами была проведена гибридизация картофеля в условиях горного пояса нашей республики для получения гибридов картофеля и изучения особенности проявления сил гетерозиса у гибридов.
Для определения всхожести семян сортов и гибридов картофеля, был заложен опыт в лабораторных условиях, а также в марлевом изоляторе в Файзабадском районе.
Для подсчёта количества семян и проведения посева, использована разработанная нами методика «свернутой бумаги». Этот метод прост и удобен для проведения подсчёта мелких семян картофеля, легко и быстро осуществим. Для посева использовали пластиковые сосуды длиной 20см, шириной и высотой соответственно 10х10см, субстрат из смеси перепревшего навоза, почвы и песка в соотношения 3:1:1. Сначала сосуды с субстратом поливались лейкой. Затем в каждый сосуд высевали по 50 шт. семян, поверхность которых укрывали перепревшим навозом, на 0,5 -0,7 см. Сосуды поливали через каждые двое суток, обеспечивая тем самым повышенную влажность субстрата с посеянными семенами (рисунок Первые всходы семян картофеля появились через 7-10 дней после посева.
Сорта и гибриды картофеля на 15 – й день от посева, имели разное количество всходов. Например, семена сорта Зарина и ряда гибридов Клон -66 х Файзабад; Клон 48 х Кондор; Кардинал х Кондор и Кардинал х Пикассо в этот период дали всходы от 20 до 32%. Наряду с этим, такие сорта, как Пикассо, Кондор и Клон -36/6 и гибриды Клон- 48 х Дусти и Клон- 48 х Дусти дали всего лишь 2- 4% всходов.
Сорт Кардинал и гибриды ВИР-51/90-7-2, ВИР -59/190-4, ВИР -180-1, ВИР-64/90-7-7, ВИР-93-5-30 и ВИР-51/90-7-2 в этот период всходов не дали.
Такие разные показатели по всходам наблюдались между сортами и гибридами также на 20-й, 25-й и 30 – й дни после посева семян. Среди сортов на 30 – й день от посева, низкий показатель по всхожести наблюдался у сорта Кардинал (18%), а самый высокий - у сорта Зарина (62%). Среди гибридов низкий показатель всхожести семян наблюдали у гибридов ВИР-51/90-7-3, ВИР-51/90-7-2 и ВИР -180-1 (соответственно 10, 12 и 14%); высокая всхожесть семян наблюдалась у гибридов Файзобод х Пикассо, Кардинал х Пикассо, Кардинал х Пикассо, Зарина х Дусти -52- 54%.
Самую высокую всхожесть семян наблюдали у гибридной комбинации Кардинал х Кондор (84%), хотя у материнской формы данного гибрида, сорта Кардинал, всхожесть составила только18%, а у отцовской формы сорта Кондор-26%. Это, видимо, связано с эффектом силы гетерозиса у данного гибрида по данному признаку.
Таким образом, всхожесть семян сортов и гибридов картофеля при выращивании в условиях лаборатории в сосудах, тесно связана с особенностями сортов и гибридов картофеля - на 30 – й день от посева она колебалась в пределах 8 - 84%. Средняя всхожесть семян по всем сортообразцам картофеля составляла 37,2%.
Выращенные рассады сортов и гибридов картофеля через месяц были высажены в грунт в теплицу по схеме 45х 20 см, а часть их была высажена в открытом поле. Были проведены фенологические наблюдения за ростом и развитием растений. Во время уборки урожая были проведены клоновые отборы среди популяции гибридов.
В конце вегетации на основе визуальной оценки растений, при отсутствии признаков грибных, бактериальных и вирусных болезней на стеблях, листьях и клубнях, исследуя компактность гнезд, количество клубней, глубину глазков, размер столонов, окраску клубней, продуктивность кустов, легкость выделения клубней от столонов и другие признаки, провели клоновые отборы. Частота полезных клоновых отборов среди популяции гибридов F1 картофеля, составила от 4.76 до 20%
и по частоте полезных клоновых отборов, среди гибридных комбинаций, особенно отличались такие комбинации, как - F1 Файзобод х Пикассо, Клон- 48 х Дусти, Клон -23 х Пикассо, Дусти х Пикассо, Дусти х Кондор, Клон 48 х Кондор, Катадин х Зарина и Клон - 66 х Файзабад, среди популяции растений, которые имели от 10 до 20% полезных клоновых отборов. Меньшее количество клоновых отборов были выделены в таких гибридных комбинациях, как ВИР-51/90-7-2, ВИР-64/90-7-7, ВИР -59/190-4, и Клон 66 х Клон 40/, у которых число отборов составило от 4.76 до 7,0%. По остальным гибридным комбинациям клоновые отборы составили от 7.11 до 10% от общего количества популяции растений.
В среднем, количество полезных клоновых отборов среди гибридов F1 картофеля, составило 10.17% от общего количество растений в популяциях.
В первом поколении гибридов картофеля по таким качественным признакам, как окраска цветков, клубней, глазков, форма клубней, мы наблюдали в основном доминирование доминантных признаков, хотя было и расщепление по этим признакам в зависимости от типа скрещивания сортов картофеля.
Картофель является сложным объектом для генетических исследований в силу того, что многие сорта картофеля по генотипу являются гетерозиготными. Однако картофель помимо генеративных органов (семян), размножается и через вегетативные органы (клубни, ростки, почки, столоны), представляя определенный интерес для селекционно-семеноводческих работ.
Многие исследователи, проводя свои исследования по изучению характера наследования таких качественных признаков у аутотетраплоидных видов картофеля (S.tuberosum L. и другие), как окраска клубней, венчиков, ростков, глазков, форма клубней, венчиков, листьев, пришли к выводу, что многие из этих признаков контролируются разными генами (Альсмик П.И.,1979; Будин К.З., 1986; Х,Росс, 1989; А.П.,Гончаров Н.Д., 1996; Киру С.Д., 2009; Ермишин А.П., 2010).
По сообщениям (Яшина И.М. и др., 1973), к числу аутотетраплоидов относятся культурные тетраплоидные виды S. tuberosum и S. andigenum (2n=48). Многие локусы у этих видов наследуются по типу тетрасомии. Особенности расщепления у аутотетраплоидов определяются наличием четырех гомологичных хромосом, случайной их конъюгацией в мейозе и случайным распределением по гаметам. Различное число доминантных аллелей в определенном локусе в каждой из четырех гомологичных хромосом обусловливает у аутотетраплоидов появление трех типов гетерозигот (АААа, ААаа, Аааа) и двух типов гомозигот (АААА и аааа).
Эти пять возможных генотипов в соответствии с числом доминантных аллелей, входящих в их состав, называются: квадриплексом (АААА), триплексом (АААа), дуплексом (ААаа), симплексом (Аааа) и нуллиплексом (аааа). Часто используют сокращенные формулы этих генотипов: А4 , А3а, А2а2, А1а3 и а4.
У аутотетраплоидов имеются пять возможных вариантов моногибридных скрещиваний, дающих расщепляющиеся по фенотипу потомства (в отношениях 35:1, 11:1, 5:1,3:1 и 1:1), в то время как у диплоидов только два варианта моногибридных скрещиваний (Аа Х Аа и Аа Х аа), которые приводят к расщеплению в отношениях 3:1 и 1:1.
Экспериментальные данные о дигибридном и тригибридном расщеплениях у сортов картофеля очень ограничены, поскольку для проведения подобных исследований необходим анализ большого количества потомков. Имеющиеся экспериментальные материалы относятся только к тем случаям, когда изучаемый признак контролируется совместным действием двух генов (комплементарное взаимодействие и эпистаз).
Недостаток, типичный для автотетраплоидов, - это трудность прослеживания генетики различных признаков (Росс Х., 1989).
У селекционных сортов комплементарно наследуются такие хозяйственно-важные признаки, как устойчивость к раку, совместное действие генов Y и Z ( Lunden A.P., 1937), устойчивость к вирусу X, совместное действие генов A и B (Stevenson F.J. et al., 1939), все типы окраски клубневой кожуры, совместное действие генов RE или RD, контролирующих красную окраску клубней, и совместное действие генов PE или PD, контролирующих синюю окраску (Т.В. Ассеева и др., 1935; Lunden A.P., 1937).
По данным авторов (Т.В. Ассеева и др., 1935; Lunden A.P., 1937), окраска цветков картофеля также является комплементарно - наследуемым признаком, и определяется совместным действием генов PF (сине-фиолетовые цветки) или RF (красно-фиолетовые цветки). Эпистатическое взаимодействие имеют гены P и R, определяющие совместно с геном D или E соответственно, синюю или красную окраску клубней, а совместно с геном F – аналогичную окраску цветков. Здесь наблюдается одновременное комплементарное и эпистатическое взаимодействие генов, и фенотипическое отношение в потомстве, рассчитанные по формуле (P : p) X (R : r) X (D :d) или (P : p) X (R : r) X (E: e), которое будет зависеть как от дозы каждого гена в генотипе родительских форм в симплексном, дуплексном или других состояниях, так и от взаимодействия между генами и их фенотипическими эффектами.
Как сообщает (Брюбейкер Дж.Л., 1966), наследование количественных признаков у аутотетраплоидов еще сложнее по сравнению с диплоидами. Число образующихся фенотипов при расщеплении по количественному признаку, в развитии которого участвуют пять диаллельных локусов, может дать в потомстве дуплекса 55 = 3125 фенотипов против 35= 243 в потомстве диплоидной гетерозиготы. У картофеля (Яшина И.М. и др.,1973) отбор по количественным признакам облегчается благодаря вегетативному размножению, позволяющему подхватывать фенотипы с максимальным проявлением признака, в том числе и гетерозиготные.
По сообщениям (Яшина И.М. и др., 1973), фенотипически различают белую, красную, (розовую) и синюю окраску клубней; белую, красно-фиолетовую, сине-фиолетовую и синюю окраску цветков и аналогичную им окраску ростков, а также красноокрашенную, синеокрашенную и неокрашенную окраску глазков.
Е.К. Эмме (1936) установила, что окраска венчика обусловлена взаимодействием нескольких генов (Ansp , Aninf, Cr, Cy), из которых Cr и Cy по– видимому, представлены множественными аллелями, дающими широкий спектр интенсивности окрасок. Как правило, синяя окраска доминирует над красной и белой, а красная – над белой.
Английскими исследователями (Dodds K.S. et. al., 1956) при скрещивании клонов S. rybinii C.P.C. 979 с синими цветками и S. rybinii C.P.C. 2211 с белыми цветками, в F1 обнаружили выщепление синецветковых (125) и красноцветковых (113) растений в отношении 1:1. При беккроссах синецветковых гибридов F1 на C.P.C. 2211 (белые цветки) в B1 получали четыре класса растений с синими, голубыми, красными и белыми цветками (соотношение 1:1:1:1), а при беккроссах красноцветковых гибридов F1 – два класса растений с красными и белыми цветками (1:1), что свидетельствовало о бифакторальной природе окраски венчика.
Е.К. Эмме (1937) при скрещивании белоклубневой формы S. rybinii с желтоклубневой формой S. Goniocalyx, в потомстве наблюдала расщепление в отношение трех желтоклубневых на один белоклубневый сеянец (33:13), свидетельствующий о доминировании желтой окраски клубней над белой.
Как сообщают Dodds K.S. и др., (1956), основная окраска клубней контролируется тремя независимыми локусами – P, R (с серией множественных аллелей) и I, из которых первые два участвуют в генетическом контроле окраски венчика. Эпистатичный аллель i (локуса I) в гомозиготном состоянии обусловливает отсутствие окраски клубня.
Сообщается (East E.M., 1910, Salaman R.N., 1910), что у диплоидных видов картофеля синяя окраска клубней доминирует над красной, и оба они доминируют над белой. По информации Salaman R.N. (1910), окраску клубней контролируют три независимые доминантные гены: P – синяя окраска, R- красная окраска и D- комплементарный ген. Комбинация генов PRD дает синие клубни, а DR – красные. Растения с одним каким-либо геном (P, R или D) имеют белые клубни, как рецессивы prd.
По результатам анализа комбинации Любимец Х Альма (Яшина И.М. и др., 1973) из 148 проанализированных растений у 35 были красные клубни, а у 113 белые клубни, что близко к отношению 1:3.
Нами в течение 2010-2012 гг. изучен характер наследования таких качественных признаков у гибридов картофеля F1, как окраска цветков, клубней и глазков в условиях горной зоны Таджикистана.
Качественные основные признаки родительских сортообразцов картофеля, которые были изучены в опытах по гибридизации картофеля,
Полученные данные по характеру расщепления качественных признаков у гибридов картофеля F1 показали сложную природу этого явления среди популяций гибридов (табл. 12).
Таблица 12. Расщепление гибридов картофеля F1 по качественным признакам
|
Гибридные комбинации |
Число растений |
Растение с окраской цветков: |
Растение с окраской клубней: |
Растение с окраской глазков: |
||||||
|
фиолетовая |
белая |
отношение |
желтая |
красная |
отношение |
неокрашенная |
красная |
соотношение |
||
|
Дусти х Пикассо |
84 |
0 |
84 |
0 |
84 |
0 |
0 |
70 |
14 |
4,7:1 |
|
Файзабад х Пикассо |
69 |
0 |
69 |
0 |
69 |
0 |
0 |
54 |
15 |
3,6:1 |
|
Кардинал х Пикассо |
77 |
55 |
22 |
2,5:1 |
53 |
24 |
2,2:1 |
60 |
17 |
3,5:1 |
|
Зарина х Дусти |
78 |
58 |
20 |
2,9:1 |
78 |
0 |
0 |
78 |
0 |
0 |
|
Клон 23 х Пикассо |
70 |
49 |
21 |
2,3:1 |
51 |
19 |
2,7:1 |
56 |
14 |
4,0:1 |
|
Кардинал х Кондор |
78 |
78 |
0 |
0 |
0 |
78 |
0 |
78 |
0 |
0 |
|
Дусти х Кондор |
89 |
65 |
24 |
2,7:1 |
63 |
26 |
2,4:1 |
89 |
0 |
0 |
Как видно из табл.12, гибриды картофеля F1, в зависимости от вида гибридной комбинации, по признаку окраски цветков расщепляются на два класса – растения с фиолетовой и белой окраской цветков. Фиолетовая окраска цветков, по отношению к белой окраске, является доминантной и отношение между этими признаками составляет 2,3- 2,9:1.
Гибриды также в зависимости от признаков родителей расщепляются на два класса по признаку окраски клубней. По данному признаку наблюдается расщепление гибридов в соотношение 2,2-2,7:1. По признаку окраски глазков у гибридов наблюдается расщепление по двум классам растений – с неокрашенными и с красными глазками. По данному признаку расщепление наблюдается в отношении 3,5 - 4,7:1.
На основе полученных нами данных, можно предполагать, что такие признаки, как фиолетовая окраска цветков по отношению к белой, желтая окраска клубней по отношению к красной и неокрашенность глазков по отношению к красной окраске глазков, проявляются у гибридов картофеля F1 как доминантный признак. Степень проявления этих отношений зависит от генетической природы этих признаков у родительских форм и вида гибридной комбинации.
Особенно высокое отношение доминантного признака неокрашенности глазков по отношению к красным глазкам, наблюдается у гибридных комбинаций Дусти х Пикассо и Клон 23 х Пикассо (соответственно 4.7:1 и 4.0:1).
Выделенные клоны среди популяции гибридов F1, были изучены в F1 С1 (первое клубневое поколение или питомник изучения гибридов первого года) в сравнении с родительскими формами в 2011 году в условиях Джиргитальского района, на высоте 2700 м н.ур. моря.
По признаку клубней на одном растении, лучшие показатели имеют клоны 6 tj; 12 tj; 14 tj; 16 tj; 17 tj; 22 tj; 23 tj; 29 tj и 32 tj, у которых насчитывается от 10 до 18 шт. клубней, что почти в два – три раза больше, чем у других клонов. По весовому признаку клубней, особенно отличались клоны 1 tj; 7 tj; 10 tj; 13 tj; 18 tj; 20 tj; 27 tj; 31 tj; 37 tj; 40 tj, ВИР-93 и другие. Масса одного клубня у этих клонов колебалась в пределах от 90 до 218 г, что по сравнению с другими клонами, больше в полтора - два раза.
По признаку продуктивности клонов особенно отличались клоны 1 tj; 10 tj; 14 tj; 23 tj; 29 tj; 31 tj; 32v; 34 tj; 40 tj; 53 tj и другие. У этих клонов продуктивность одного растения составила более 800 – 1000 г, что на 40 - 70% больше, чем у других клонов картофеля.
Полученные новые гибриды картофеля представляют большой интерес для селекции картофеля в Таджикистане. Клубни перспективных клонов переданы для оздоровления и ускоренного размножения методами биотехнологии в лаборатории молекулярной биологии и биотехнологии.
Данные, полученные нами по определению гипотетического гетерозиса у гибридов F1C2 картофеля, приведены в табл.13.
Таблица 13 Гипотетический гетерозис у гибридов F1C1 картофеля, % (Джиргитальский район, 2011г.).
|
Гибридная комбинация |
по количеству клубней. |
По массе 1-го клубня |
по продуктивности |
|
(Дусти x Кондор) |
47.40 ±1.7 |
35.25 ±2.7 |
25.68±4.1 |
|
(Файзабад x Пикассо) |
84.57±1.9 |
20.0±2.6 |
20.15±3.9 |
|
(Клон-23 x Пикассо) |
58.62±2.7 |
26.62 ±2.3 |
59.01±4.1 |
|
(Зарина x Дусти) |
12.5±2.2 |
29.16±2.5 |
23.56 ±4.1 |
|
(Дусти x Пикассо) |
59.16±2.8 |
41.15±2.2 |
73.01±5.1 |
|
(Кардинал x Пикассо) |
18.95 ±2.6 |
40.05±2.5 |
26.77±4.6 |
|
(Клон - 48 x Кондор) |
81.20±2.4 |
73.57±2.9 |
49.75±4.8 |
|
(Кардинал x Кондор) |
71.53±2.1 |
36.07±2.3 |
52.34±4.9 |
|
(Клон-66 x Файзабад) |
73.33 ±2.7 |
42.54±2.9 |
76.59±5.2 |
|
(Клон 40/1x Дусти) |
20.12±3.1 |
30.04 ±2.8 |
14.04±4.3 |
|
Среднее |
55.85 |
37.44 |
42.08 |
|
V,% |
25.81 |
18.97 |
21.45 |
|
НСР 05 |
18.10 |
13.25 |
14.37 |
Из табл.13 видно, что гипотетический гетерозис у гибридов по признаку количества клубней на одно растение (на одном растении), колеблется от 12.5 до 84.57%. Самый высокий показатель по этому признаку, наблюдался у гибрида F1C1 - Файзабад x Пикассо – 84.57%, а самый низкий – по гибриду F1C2 -Зарина x Дусти – 12.5%.
Средний показатель по гипотетическому гетерозису по всем гибридам составил 55.85%.
По весовому признаку клубня, эффект гетерозиса сильно проявляется у гибридов из комбинации F1C1 - Клон - 48 x Кондор, Дусти x Пикассо и Клон-66 x Файзабад, у которых этот показатель составляет более 41-73%.
Однако, у таких гибридов, как Файзабад x Пикассо, Клон 40/1x Дусти, Зарина x Дусти и Клон-23 x Пикассо эффект гетерозиса составляет 20-30%. А средний показатель данного признака составил 37,44%.
Гипотетический гетерозис у гибридов картофеля, в целом по признаку продуктивности растений, варьируется в пределе 14 – 73%.
Наиболее высокий показатель по данному признаку наблюдается по таким гибридным комбинациям, как Клон-66 x Файзабад и Дусти x Пикассо, у которых этот вид гетерозиса составляет более 70%, тогда как по другим гибридам он составляет 14 - 59%.
В среднем гипотетический гетерозис по продуктивности составляет 42.08%.
Таким образом, у гибридов F1C1 картофеля наблюдается большой диапазон распределения показателей по гипотетическому гетерозису по таким признакам как, количество клубней, масса одного клубня и продуктивность растений. Высокий коэффициент доминирования наблюдался также по признаку продуктивности растений
Коэффициент доминирования признака продуктивности растений бывает низким (0.42-1.47) у гибридов F1C1 - Клон 40/1x Дусти, Дусти х Кондо и Клон - 48 x Кондор. Однако гибриды F1C1 - Кардинал х Кондор, Клон-23 x Пикассо и Дусти х Пикассо имеют высокие показатели по этому признаку (5.47-28.82). В среднем коэффициент доминирования признака продуктивности у гибридов F1C1 картофеля составил 8.13.
Как известно, продуктивность растений картофеля в основном зависит от числа клубней и среднего веса клубня. Как сообщают W.A. Riedl (1948) и K.H. Muller (1965), эти признаки передаются по наследству. K.H.Engel (1957) установил высокую положительную корреляцию между средним весом клубней и урожаем первого поколения, и сообщил, что величина клубней является более постоянным признаком, чем их число.
Результаты скрещивания, проведенные И.М. Яшиной и др. (1973) показали, что по урожайности у картофеля наблюдается гетерозис. Урожай растений в самоопыленном потомстве всегда ниже: гомозиготность ведет к деградации потомства, угнетению. При скрещивании, потомство имеет более высокую урожайность, что связано с более высокой степенью гетерозиготности, т.е. гетерозисом.
По сообщениям F.A.Krantz (1925), направленный инбридинг и скрещивание инбридных (инбридинговых) клонов между собой, приводит к получению высокоурожайного потомства. Гибридные семьи, полученные от скрещивания сорт х сорт, сорт х инбред и инбред х инбред, не отличаются по уровню средней урожайности. Однако в потомстве инбред х сорт наблюдался более высокий процент высокоурожайных гибридов по сравнению с потомством инбред х инбред.
Как указывают R. Schick и др. (1962), оценку комбинаций по урожайности можно сделать только в первой клубневой репродукции, так как по урожайности сеянцев из семян, нельзя судить об истинной урожайности гибридов. Однако как сообщают Семенова И.А. (1966) и Гончаров Н.Д. (1996), достоверную оценку урожая можно провести не раньше, чем во второй клубневой репродукции, когда величина урожая и размер клубней проявляются в полной мере.
Как известно, продуктивность растений картофеля в основном зависит от числа клубней и его среднего веса. Как сообщают (Riedl W.A., 1948; Engel K.H., 1957; Семенова И.А., 1965; Яшина И.М. и др.,1973; Альсмик П.И., 1979), эти признаки передаются по наследству, и они имеют высокую положительную корреляцию с продуктивностью к другим признакам (от r = 0.012 до r = 0.586).
Исследования показали, что между признаками количества клубней и продуктивностью, наблюдается положительная средняя корреляция у гибридов F1C1 (в первом клубневом поколении) картофеля.
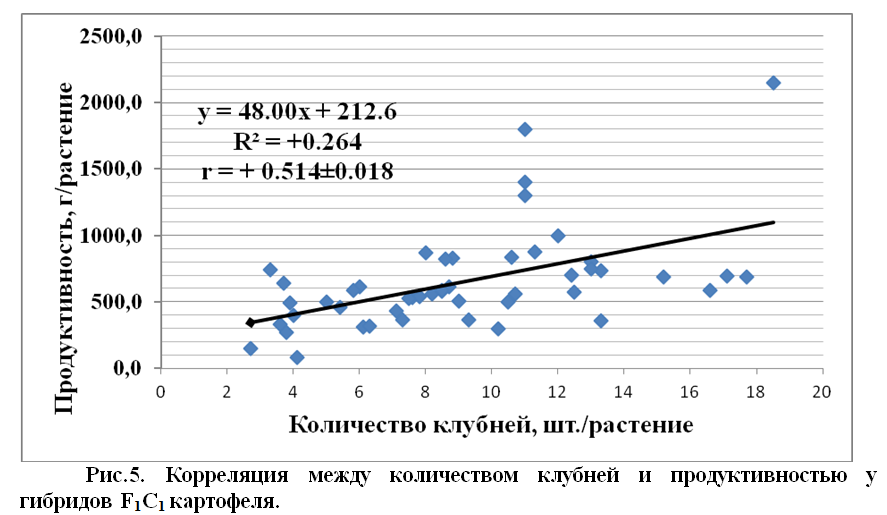
В среднем, коэффициент корреляции между признаками количества клубней и продуктивности у гибридов, составляет r = 0,514±0.018, что можно отнести к среднему уровню положительной корреляции. Коррелятивная связь этих признаков у родительских форм равнялась к r = 0,912±0.011, и она была значительно выше, чем у гибридов F1C1.
По сообщениям Абдуллаева Х.А. и др. (1990), характерной особенностью наследования количественных признаков, является их взаимная корреляция, что относится и к количественным признакам фотосинтеза.
Авторы утверждают, что тесная связь признаков обусловлена, с одной стороны, сцеплением полигенов или их плейотропным эффектом (генетическая корреляция), с другой, – влиянием агроклиматических факторов, когда адаптационное изменение одного признака под влиянием условий внешней среды может вызвать альтерацию других признаков (паратипическая корреляция).
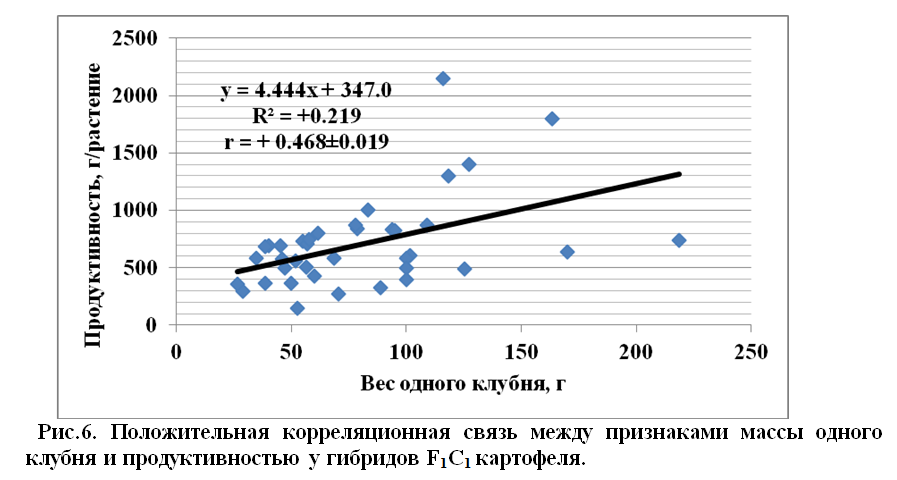
Коррелятивная связь между массой одного клубня и продуктивностью растений у гибридов F1C1 картофеля составила r = + 0,468±0.019 (рис.6). Этот показатель у родительских форм составил r = + 0,881±0.017.
Как сообщает А.Ф. Салимов (2007), корреляционный анализ выявил прямую положительную связь урожайности сухой фитомассы и клубней с параметрами агроценоза картофеля. Наиболее сильная, она у урожая с сухой фитомассой, с площадью листьев (r = 0,997), у урожая клубней с ЧПФ (r = 0,995), у урожая от массы микроклубней r =(0,835), и от (у) схемы посадок (r = 0,938).
В наших опытах между признаками количества клубней и массой одного клубня, корреляция оказался отрицательной r = - 0,335±0.022, не только у гибридов F1C2 , но и у родителей r = - 0,251±0.024 (рис.7).
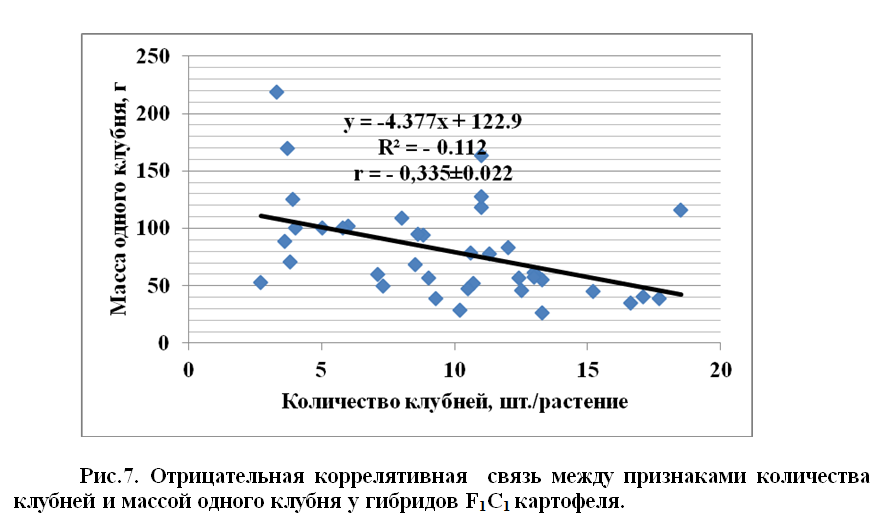
Установлено что, между признаками количества клубней, массой одного клубня и продуктивности растений, существует положительная коррелятивная связь, а между количеством клубней на одно растение и массой одного клубня – отрицательна, у гибридов F1клубневое поколение) (Клон-48 x Кондор) выделен клон, который имел 130 шт. ягод, вес которых составил 1200 г. Извлеченных семян у этого растения оказалось 21058 шт.
Кроме того, данный клон имел 21 шт. клубней с массой 1500 грамм. Такие клоны сохранены, и в перспективе будут использованы для скрещивания и закрепления признака многоплодовитости (многоплодности) (формирования большого количества ягод).
Также в своих работах о большом разнообразии и формировании цветков и ягодообразования у картофеля, сообщают Подгаецкий А.А. и др., 2008.
Среди гибридов картофеля наблюдается также большое разнообразие признаков окраски и формы клубней и глазков.
В вегетационный период 2012 года, в условиях Джиргитальского района, определяли продуктивность гибридов F1С2 картофеля (табл.14).
Таблица 14 Продуктивность гибридов F1 C2 картофеля в условиях Джиргитальского района (2012г.).
|
№ |
Номера клонов по каталогу ИБФГР АН РТ |
Гибриды картофеля |
Кол. во растений, шт. |
Продуктивность, г/растение |
Отклонение от стандарта по хозяйственной продуктивности, % |
|
|
биологическая |
хозяйственная |
|||||
|
1 |
- |
Кардинал (стандарт) |
55 |
745 |
609 |
100,0 |
|
2 |
ВИР-93 |
ВИР-93 |
57 |
1100 |
800 |
131,4 |
|
3 |
Клон-2tj |
Дусти x Кондор |
56 |
1574 |
1310 |
215,1 |
|
4 |
Клон-9 tj |
Дусти х Кондор |
55 |
1240 |
900 |
147,8 |
|
5 |
Клон-14tj |
Дусти х Кондор |
57 |
905 |
800 |
131,4 |
|
6 |
Клон- 23tj |
Гибрид -23 x Пикассо |
58 |
1800 |
1365 |
224,1 |
|
7 |
Клон-33tj |
Дусти х Кондор |
56 |
2200 |
1420 |
233,2 |
|
8 |
Клон -40 tj |
Клон-66 x Файзабад |
53 |
1698 |
1132 |
185,9 |
|
9 |
Клон- 44tj |
Клон-36/6 |
56 |
1460 |
800 |
131,4 |
|
10 |
Клон- 51tj |
Гибрид -23 x Пикассо |
57 |
1500 |
850 |
139,6 |
|
11 |
Клон-53 tj |
Кардинал х Кондор |
56 |
1800 |
1300 |
213,5 |
|
|
|
Среднее |
|
1456,8 |
1026,0 |
168,5 |
|
|
|
НСР05 |
- |
70,5 |
51,7 |
- |
|
|
|
|
|
|
|
|
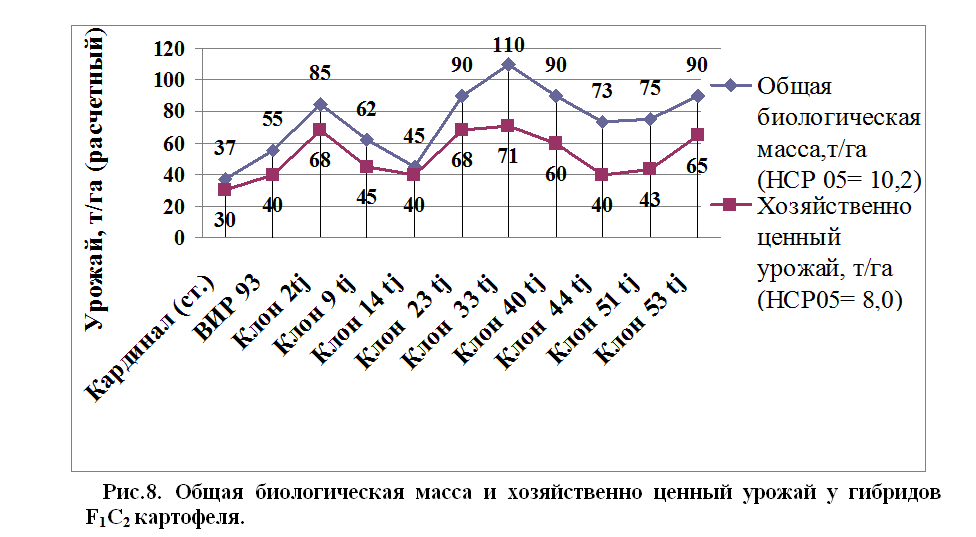
Из табл.14 видно, что новые гибриды F1 C2 картофеля, как по общей продуктивности (биомассе), так и по хозяйственно ценной (клубней), значительно превосходят стандартный сорт картофеля Кардинал. По общей продуктивности особенно отличаются такие клоны, как Клон-2tj; Клон- 23tj; Клон-33tj; Клон-53 tj, которые по общей и хозяйственной продуктивности превышают сорт Кардинал в полтора - два раза. Отобранные клоны имеют полезные показатели по признакам количества клубней, массе одного клубня и продуктивности. Коэффициент вариации по этим признакам варьируется от 43 до 94%, что свидетельствует о большой изменчивости в разрезе гибридных комбинаций.
По признаку расчетной урожайности, почти все выделенные новые гибриды картофеля, превышают сорт Кардинал. В частности, общий урожай биомассы у гибридов F1С2 картофеля колеблется от 45 до 110 т\га, тогда как этот показатель у сорта Кардинал составляет 37 т\га. Такая картина наблюдается и по признаку хозяйственного урожая (урожая клубней) гибридов F1С2. У гибридов картофеля хозяйственный урожай колеблется в диапазоне – 40-71 т/га, а у сорта Кардинал он составляет 30т/га, что на 31-133% меньше по сравнению с гибридами F1С2 картофеля.
Полученные новые гибриды картофеля имеют большой потенциал накопления биологического и хозяйственного урожая, что является важным фактором для получения урожая клубней, как продукта питания, так и столь необходимой биомассы в виде остатков стеблей и корней для обогащения почвы органикой. Необходимо отметить, что эти гибриды картофеля накапливают значительно большее количество биомассы при одинаковых условиях возделывания со стандартным сортом Кардинал (одинаковые дозы внесения минеральных удобрений и выполнения, идентичных агроприемов возделывания). По признаку соотношения хозяйственно ценного урожая клубней к общей биологической массы, самый высокий показатель наблюдается у Клона -14tj (89%).
Таким образом, путем использования методов классической селекции (гибридизация и визуальный клоновый отбор) получены новые гибриды картофеля, которые представляют большой научно - практический интерес для дальнейших селекционно - семеноводческих работ.
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()