АгроСборник.Ру

С.Ф. Коваль, В.П. Шаманин "Растение в опыте"
При нарушении оттока интенсивность фотосинтеза некоторое время может поддерживаться депонированием продуктов в вакуолях, в эндоплазматическом ретикулуме или межклеточном пространстве. Возможно отложение их в паренхиме листовых черешков и стебля. Именно благодаря этому возможно корректное определение фотосинтеза на высечках листьев и отделенных органов. Быстрый отток ассимилятов из листа восстанавливает фотосинтез. Если внешние условия не лимитируют его, то ведущая роль в детерминации валовой продукции принадлежит акцепторам фотоассимилятов. Появление новых аттрагирующих центров (точек роста, запасающих органов), а также частичная потеря листовой площади увеличивают интенсивность фотосинтеза [Мокроносов, 1970]. При этом, вначале происходит повышение интенсивности фотосинтеза существующей листовой поверхности, а позднее - и увеличение валового фотосинтеза в результате ускорения роста.
Физиология аттракции почти не изучена, но известно, что в основе ее лежат компетентность потребляющих органов (готовность к выполнению функции), связанная с их гормональным режимом. Действие гормонов и системе растительного организма изложено в сводках (Дерфлинг, 1985; Мокроносов, 1986; Полевой, 1982; Кефели, 1974]. В опытах удается усилить приток метаболитов к тем или иным органам путем местной обработки гормонами (индолилуксусная кислота, кинетин и др.). Изучение силы аттракции основано на контроле распределения между органами радиоактивной метки или на наблюдениях за изменением роста, накопления сухого веса после удаления некоторой части акцепторов метаболитов [Пинхасов, 1981].
Удаление колосков с одной стороны колоса (пиницировка) приводит к увеличению среднего веса зерна в оставшихся колосках, если их налив лимитировал фонд метаболитов. Если налив зерна на интактном растении лимитировался аттрагирующей силой колоса, то пиницировка, уменьшающая его суммарную аттракцию приводит к снижению среднего веса зерна в оставшихся колосках [Коновалов, 1981].
В молодом растении весь фонд вновь образовавшихся ассимилятов используется на формирование новых производящих органов (листьев, корней). В результате продуктивность нетто-фотосинтеза во времени возрастает по правилу сложных процентов - каждая новая единица живой биомассы создает новую производящую биомассу растения. С переходом и генеративную фазу или с началом формирования запасающих органов часть ассимилятов отвлекается новыми акцепторами, нарастание производящих органов тормозится. Распределение общего фонда метаболитов между этими потребителями зависит от аттрагирующей силы соответствующих органов. Кроме того, во все фазы онтогенза часть наполнения сахарами ранее образованных клеток и тем замкнуть виток обратных связей: фотосинтез —> спрос на ассимиляты быстрое потребление в акцепторе —> стимуляция фотосинтеза —> рост потребления.
Время от времени появляются сообщения о достижении селекционными или агротехническими методами рекордного размера плодов у какой-то культуры. Далее начинаются спекулятивные расчеты возможного в связи с этим увеличения урожая. На деле же, такие сообщения имеют интерес только при указании числа плодов в сравнении со стандартными сортами (как на растение, так и на единицу площади посева).
Наличие многих акцепторов ассимилятов создает дробление их потока и образование многих малопродуктивных органов. Преобладание аттракции одного или немногих из них над остальными даст в результате крупные, но малочисленные плоды. Так отбор томатов на крупность плода одновременно ведет к уменьшению их числа при сохранении той же хозяйственной продуктивности растения. Без существенного изменения общего фонда метаболитов реализуемые сочетания крупности плодов (семян) и их числа на растении располагаются вдоль линии регрессии число- крупность. Такие регрессии являются параболами, совпадающими с линиями неизменной хозяйственной продуктивности (Рис. 3.3).
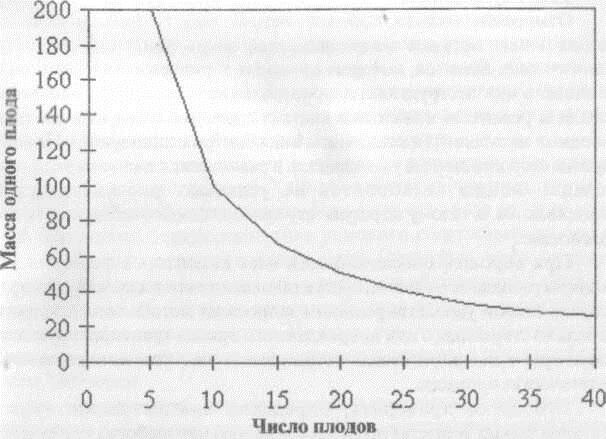
Рис. 3.3. Зависимость веса одного плода от их числа на растении томата.
Разрешение этого противоречия возможно следующими путями. 1. Увеличением фонда метаболитов, транспортируемых в хозяйственно ценные органы. 2. Уменьшением числа акцепторов при одновременном максимальном усилении а прагирующей силы у тех из них, которые остались на довольствии. Первое связано с улучшением экологических условий и быстрым оттоком ассимилятов из хлоропластов в органы-акцепторы. Второе
томата с «высоким запросом» акцепторов на метаболиты и точным адресом их транспортировки. В итоге будет иметь место не только увеличение общей биологической продуктивности растения, но и доли биомассы в хозяйственно ценных органах - т.е. увеличение К/хоз.
Сейчас признается, что биологическая продуктивность примитивных и современных сортов практически одинакова. Все успехи селекции за последнее столетие связаны с увеличением доли биомассы, сосредоточенной в хозяйственно ценных органах [Батыгин, 1986]. Таким образом, рост хозяйственной продуктивности имеет причиной не увеличение нетто-фотосинтеза, а увеличение К/хоз [Khalifa, Al-Saheal, 1984; Семенов и др., 1985; Takeda, Frey, 1985; Герасименко, 1988] в результате перераспределения аттракции между органами. Малое влияние экологических условий на этот показатель [Кадыров и др., 1984; Sharma et al., 1984] делает его весьма удобным параметром для отбора на ранних этапах селекции.
Поделиться!
Понравилась статья? Расскажите о ней знакомым или оставьте комментарий!
![]()
![]()
![]()
![]()
![]()
![]()
![]()